


اطلاعات عمومی,ویکی ژن
ماکرولیدها (Macrolide): نحوه فعالیت و مکانیسم مقاومت در برابر آنها
مقدمهای بر ماکرولیدها
ماکرولیدها عمدتاً عوامل ضد باکتریایی باکتریواستاتیک (Bacteriostatic) هستند که با اتصال به زیر واحد 50S مربوط به ریبوزوم، سنتز پروتئین باکتریایی را مهار میکنند. اعضای گروه شامل اریترومایسین (Erythromycin)، آزیترومایسین (Azithromycin)، کلاریترومایسین (Clarithromycin)، روکسیترومایسین (Roxithromycin)، دیریترومایسین (Dirithromycin) و تلیترومایسین (Telithromycin) هستند.
ماکرولیدها در اصل از Saccharopolyspora erythraea (که در ابتدا Streptomyces erythreus نامیده میشد) که یک نوع باکتری خاکزاد (Soil-borne) است، مشتق شدهاند.
اریترومایسین (Erythromycin) اولین ماکرولیدی بود که کشف شد و اولین بار در سال 1952 به عنوان جایگزین پنیسیلین (Penicillin) در مواردی که بیماران به پنیسیلین حساسیت داشتند یا بیماریهای مقاوم به آن داشتند استفاده شد.
ماکرولیدها در برابر باکتریهای گرم مثبت (به استثنای انتروکوک (Enterococci)) موثر هستند اما در برابر اکثر جنسهای باکتریای گرم منفی به دلیل مشکلات جذب مرتبط با غشاهای بیرونی باکتریهای گرم منفی موثر نیستند.
آنها همچنین در برابر مایکوپلاسما پنومونیه (Mycoplasma pneumoniae)، ترپونما پالیدوم (Treponema pallidum)، بوردتلا پرتوسیس (Bordetella pertussis)، کلامیدیا تراکوماتیس (Chlamydia trachomatis)، کلامیدوفیلا پنومونیه (Chlamydophila pneumoniae)، گونههای لژیونلا (Legionella)، گونههای کمپیلوباکتر (Campylobacter) و گونههای بورلیا (Borrelia) فعال هستند.
ساختار
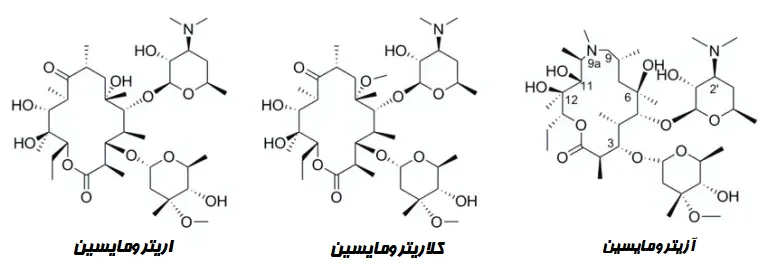
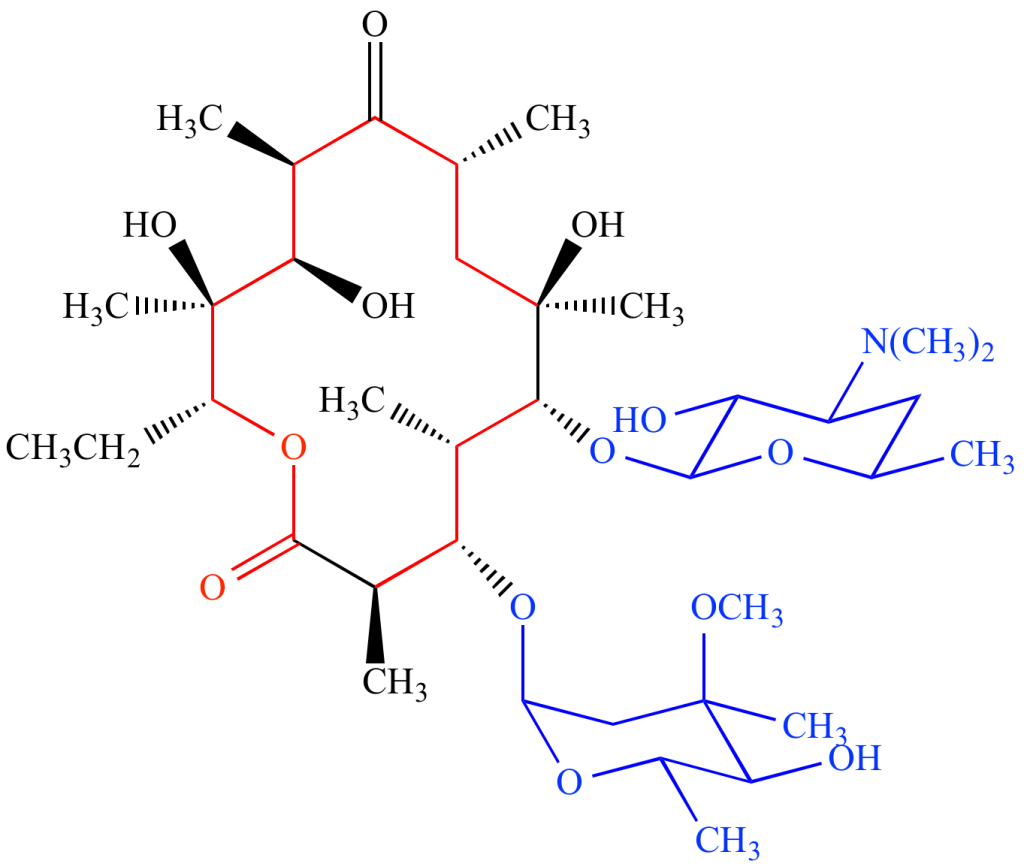
از نظر ساختاری، ماکرولیدها از یک حلقه لاکتون ماکروسیکلیک (Macrocyclic lactone) بزرگ تشکیل شدهاند که یک یا چند قند دیاکسی (Deoxy sugar) یا باقیماندههای (Residue) قند آمینهدار (Amino sugar) به آن متصل هستند.
آنتیبیوتیکهای ماکرولید بر اساس اندازه حلقه لاکتون ماکروسیکلیک (حلقه 12، 14، 15 یا 16 عضوی) و منابع (طبیعی یا سنتتیک (Synthetic)) طبقهبندی میشوند.
نحوه فعالیت ماکرولیدها
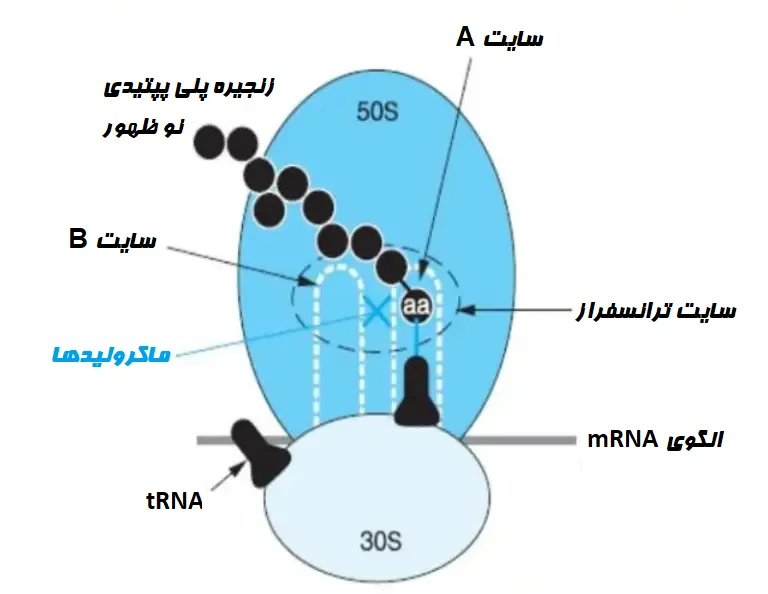
ماکرولیدها با یک هدف اختصاصی در مولکول RNA ریبوزومی 23s و پروتئینهای ریبوزومی مختلف به زیر واحد ریبوزومی 50S متصل میشوند. همه ماکرولیدها سنتز پروتئین باکتریایی را مهار میکنند، با این وجود ماکرولیدهای مختلف در مراحل متفاوتی فعالیت میکنند.
ماکرولیدهای 14 عضوی مانع انتقال پپتیدیل-tRNA (Peptidyl-tRNA) میشوند و ترکیبات 16 عضوی واکنش انتقال پپتیدیل را مهار میکنند. جدیدترین فرضیه بیان میکند که همه ماکرولیدها جدایی پپتیدیل-tRNA را از ریبوزومها در طول فاز افزایش طول، تحریک میکنند که این امر منجر به مهار سنتز پروتئین میشود.
مکانیسم مقاومت در برابر ماکرولیدها
مقاومت در برابر ماکرولیدها با دو مکانیسم مختلف رخ میدهد:
هدف تغییر یافته
تغییر آنزیمی هدف ریبوزومی، اتصال دارو را کاهش میدهد.
جهش در پروتئینهای ریبوزومی 23S rRNA، L4 و/یا L22 میتواند نسبت به ماکرولید مقاومت ایجاد کند، زیرا جهش از نظر فنی در ژن 23S rRNA اتفاق میافتد. یک مکانیسم اصلی و گسترده مقاومت به کلاس ماکرولید مربوط به آنتیبیوتیکها با واسطه ژنهای erm است که متیلترانسفرازهای rRNA (rRNA methyltransferase) را کد میکنند و یک یا دو گروه متیل را به گروه آمینو اگزوسیکلیک (Exocyclic amino) واقع در 23S rRNA اضافه میکنند.
سم زدایی آنزیمی این دارو
ماکرولیدها را میتوان توسط آنزیمهایی مانند استرازها (Esterase) یا فسفوترانسفرازها (Phosphotransferase) یا گلیکوزیل ترانسفرازها (Glycosyltransferase) یا فرمیل ردوکتازهای (Formyl reductase) کدگذاری شده در پلاسمیدهای باکتریهای گرم منفی و گرم مثبت، غیرفعال کرد.
جریان فعال این دارو
دو خانواده از پمپهای خروجی ماکرولیدی وجود دارند که با واسطه رونویسی انجام میدهند:
- mef: که یک پمپ ابرخانواده تسهیلکننده اصلی است و باعث ایجاد مقاومت به اکثر ماکرولیدهای 14 و 15 عضوی میشود.
- msr: که عضوی از ابرخانواده کاست اتصال ATP (ATP-binding cassette (ABC)) و به طور کلی در برابر ماکرولیدهای 14 و 15 عضوی و استرپتوگرامین B (Streptogramin B) موجب بروز مقاومت میشود و در برابر کتولیدها (ketolide) مقاومت در سطح پایین ایجاد میکند.
همچنین بخوانید:
- مایکوپلاسما چیست؟
- تفاوت باکتری گرم مثبت و گرم منفی: 31 تفاوت با مثال
- سنتز پروتئين چگونه است؟
- ریبوزوم: تعریف، ساختار، عملکرد و سوالات متداول
مترجم: صادق حسینیکیا
