


دوره مهارت آموزی ژنتیک مولکولی
آزمایشگاه ژنیران برگزار کننده دوره های کارآموزی نیمه خصوصی ژنتیک مولکولی ( کارگاه ژنتیک مولکولی ) می باشد.
۱۰۰ ساعت آموزش ، ۲۱ جلسه (یک جلسه آخر امتحان)
کلاس های این دوره نیمه خصوصی (۴ تا ۸ نفره) برگزار میگردد.
با اعطای مدرک از سازمان فنی حرفهای، آزمایشگاه ژنیران-انجمن زیستشناسی ایران و ITE لندن (در صورت درخواست)
شروع مجدد دوره:
- ۳ شهریور
- ۱ مهر
- ۱۷ مهر (فشرده آخر هفتهها)
- ۴ آبان
- ۲ آذر
- ۲۷ آذر (فشرده آخر هفتهها)
- هزینه دوره: ۱۴ میلیون و ۹۰۰ هزار تومان
- هزینه دوره فشرده آخر هفتهها: ۱۷ میلیون و ۸۸۰ هزار تومان
∗در صورت ثبتنام برای سازمان فنیحرفهای ۲ میلیون تومان به مبالغ فوق افزوده میشود
برای ثبت نام کافیست فقط ۲۵ درصد هزینه کل را بپردازید. ۷۵ درصد باقی مانده را می توانید در اقساط ۳ ماهه با چک صیادی پرداخت کنید. اطلاعات بیشتر

ثبت نام ژنیران
امتحان و مدرک توسط ژنیران

ثبت نام ژنیران + فنی حرفهای
امتحان و مدرک توسط ژنیران و فنی حرفهای
سرفصل های دوره مهارت آموزی ژنتیک مولکولی
- آشنایی با اصول ایمنی زیستی در آزمایشگاه
- نحوه بافر سازی و محلول سازی
- آشنایی و کار با تجهیزات آزمایشگاه مولکولی
- استخراج اسیدهای نوکلئیک (DNA و RNA)
- روشهای سنجش غلظت و خلوص اسیدهای نوکلئیک
- طراحی پرایمر
- PCR
- PCR- RFLP
- ARMS PCR
- Gap-PCR
- Multiplex-PCR
- Tetra ARMS PCR
- الکتروفورز محصول PCR با ژل آگارز
- الکتروفورز محصول PCR با ژل آکریل آمید
- سنتز cDNA
- Real-time PCR در تحقیقات و پژوهش
- Real-time PCR در تشخیص بیماریها
- آنالیز Sanger Sequencing
The course entitled “Essential Lab Skills and Molecular Biology Techniques” at Geniran laboratory covers following topics
- Introduction to biosafety
- Buffers and solutions in molecular biology lab
- Introduction to molecular biology laboratory instruments
- Nucleic acid purification(DNA and RNA)
- Nucleic acid quantification
- Primer designing
- Polymerase chain reaction(PCR)
- PCR-RFLP
- ARMS PCR
- Gap-PCR
Multiplex-PCR
Tetra ARMS PCR - Agarose and polyacrylamide gel electrophoresis
- cDNA synthesis
- Real-time PCR for Clinical Diagnosis
- Real-time PCR for mRNA Quantitation
- Sanger sequencing analysis


توضیحات دوره مهارت آموزی ژنتیک مولکولی
امروزه شاخه ژنتیک مولکولی به یکی از مهم ترین شاخه ها در زمینه های مختلف پزشکی و زیست شناسی مبدل گشته است. چرا که حیات تمامی موجودات زنده بر پایه ی ژنتیک بوده و ژنتیک تعیین کننده ی تمام صفاتی ست که در یک موجود زنده بروز می کند.
در اهمیت این شاخه می توان گفت هرچه عمیق تر ژن ها را بشناسیم، در بررسی تمامی صفات موجودات، تشخیص بیماری های ژنتیکی، انتخاب درمان مناسب و مدیریت بیماری ها قدرتمندتر خواهیم بود.
یک بیماری ژنتیکی که در اثر تغییرات در سطح ژنوم به وجود می آید می تواند اثرات خود را در سطوح مختلف ژنومیک، ترنسکریپتومیک و پروتئومیک نشان دهد.
آزمایشگاه ژنیران با برگزاری دوره کارآموزی ژنتیک مولکولی ضمن اینکه تکنیک های ژنتیکی که هم در حوزه ی تحقیقات و هم تشخیص کاربرد دارند را به طور کامل آموزش می دهد، دانشجویان را در انجام عملی این تکنیک ها نیز توانمند می سازد.
دوره کارآموزی ژنتیک مولکولی شامل 21 جلسه علمی-عملی می باشد که در این مدت کارآموزان زمان مناسب را جهت یادگیری تمامی تکنیک های مربوطه و انجام آن ها به شکل عملی داشته و به خوبی تمامی سرفصل ها را یاد می گیرند. آزمایشگاه ژنیران در دوره های کارآموزی خود، تلاش بر آن دارد که تقسیم بندی گروه ها را بسته به شرایط افراد از نظر زمانی انجام داده و با تشکیل گروه های ۴ نفره، کیفیت آموزشی را بالا ببرد.
در این دوره دانشجویان پس از آشنایی کلی با محیط آزمایشگاه ژنتیک مولکولی، با اصول ایمنی مربوطه نیز به شکل تئوری آشنا می شوند و در جلسات بعد از آموخته های خود در رابطه با ایمنی زیستی آزمایشگاه های ژنتیک مولکولی، استفاده می کنند.
در ادامه تمامی سرفصل ها تک به تک به شکل تئوری آموزش داده شده و سپس پس از انجام تکنیک توسط استاد مربوطه، توسط هر فرد به تنهایی تکنیک مجددا تکرار می گردد. لذا با تکرار مناسب تکنیک ها دانشجویان به خوبی تمامی سرفصل را آموخته و در اتمام دوره قادر به انجام تمامی آن ها خواهند بود.
دانشجویان پس از گذراندن دوره کارآموزی ژنتیک مولکولی مدرک معتبر دریافت کرده که رزومه آنها را تقویت می کند. این مدرک اعتبار کافی جهت ارائه به دانشگاه های مختلف برای اخذ بورسیه تحصیلی را داشته و افراد می توانند با گذراندن دوره ژنتیک مولکولی، هم از نظر داشتن رزومه قوی تر و هم از نظر مهارت عملی، مصاحبه های مورد نظر را با موفقیت بیشتری پشت سر بگذارند.
حداقل ترم 5 به بالای رشته های زیست شناسی، علوم پزشکی و تمامی رشته های مرتبط
در صورتی که از رشته های غیرمرتبط هستید برای راهنمایی و مشاوره رایگان با آزمایشگاه تماس بگیرید.

برای دانلود درسنامه به صورت PDF روی تصویر بالا کلیک کنید…
درسنامه مولکولی (پیش نیاز دوره)
مقدمه ای از درون سلول ها
کوچکترین واحد زیستی در موجودات زنده سلول نامیده می شود. هر سلول از ارگان های متفاوتی همچون هسته، شبکه اندوپلاسمی زبر و صاف، میتوکندری، اسکلت سلولی و غیره تشکیل شده است.
درون هسته دنیایی است که توسط علم ژنتیک مولکولی مورد بررسی قرار می گیرد. ژنتیک مولکولی در وهله اول روابط بین مولکول های زیستی RNA و DNA و کاربرد آن ها در سنتز انواع پلی پپتید ها و پروتئین ها را مورد بررسی قرار می دهد. اطلاعات ژنتیکی ذخیره شده در کد های DNA (مولکول زیستی بسیار پایدار از نظر شیمیایی) با دقت بالا کپی شده و به سلول های دختری منتقل می شود.
نواحی از مولکول DNA که وظیفه ساخت یک مولکول RNA را برعهده دارند ژن نامیده می شوند. ژنوم یوکاریوت ها از دو بخش ژنوم هسته ای و ژنوم میتوکدریایی تشکیل می شود.
DNA هسته ای دارای چند ده هزار ژن به علاوه نواحی تکراری است در حالی که DNA میتوکندریایی از ژن های اندک (دهها تا صدها ژن) تشکیل شده اند. DNA دو رشته ای از طریق فرایند رونویسی، RNA تک رشته ای را ساخته ،RNA های پیامبر (mRNA) به ریبوزوم ها منتقل شده و به شکل گروه هایی از سه نوکلئوتید (کدون) رمزگشایی می شوند و ساختار خطی آمینواسیدی تشکیل دهنده پلی پپتید ها را می سازند.

ساختار اسید های نوکلئیک(RNA , DNA) :
DNA و RNA ساختارهایی بسیار مشابه دارند. هر دو واجد زیرواحدهای مونومری نوکلئوتید که در نهایت پلیمر RNA و DNA را می سازند هستند. این دو ماکرومولکول در موارد زیر با هم تفاوت دارند:
DNA (1 دو رشته ای است در حالیکه RNA تک رشته است.

3) DNA دارای چهار باز آلی آدنین A , تیمین ,Tسیتوزین C و گوانین G است در حالی که در RNA باز تیمین با باز یوراسیل U جایگزین شده است.

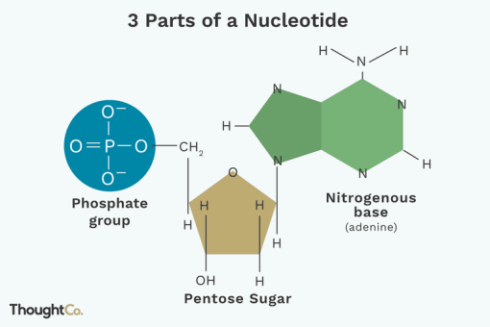
ساختار نوکلئوتید:
هر نوکلئوتید در حالت آزاد و منفرد دارای یک قند 5 کربنه ریبوز یا دئوکسی ریبوز ،یک باز آلی متصل به کربن یک پریم و سه گروه فسفات متصل به کربن 5 پریم است.
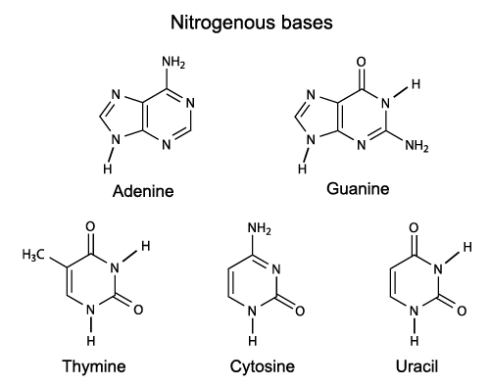
باز های آلی از حلقه های هتروسیکلیک اتم های کربن و نیتروژن تشکیل شده اند. باز های آلی به دودسته تک حلقه ای ها (پیریمیدین) و دو حلقه ای ها (پورین) تقسیم می شوند. باز های تیمین T، سیتوزین C و یوراسیل U پیریمیدین بوده و باز های آدنین A و گوانین G جزو دسته پورین ها قرار دارند.
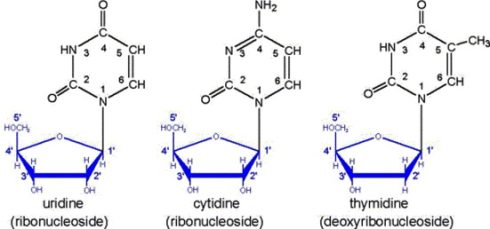
باز آلی به کربن یک پریم از قند پنج کربنه متصل می شود این ساختار را نوکلئوزید می نامند. سه گروه فسفات به کربن 5 پریم قند اتصال می یابند و ساختار نوکلئوتید را شکل می دهند. نوکلئوتید های سازنده DNA دئوکسی نوکلئوزید تری فسفات (dNTP) و نوکلئوتید های سازنده رشته RNA نوکلئوزید تری فسفات (NTP) می باشند. نوکلئوتیدها زمانی که در رشته DNA یا RNA به عنوان واحد سازنده وارد می شوند دو فسفات انتهایی خود را از دست داده و به شکل مونو فسفاته تبدیل می شوند.
با علم به اینکه مولکول های باردار در آب بسیار محلول می باشند مولکول های زیستی RNA و DNA نیز به دلیل داشتن گروه های فسفات پلی آنیون هایی با بار منفی محسوب شده و در آب حل می شوند.
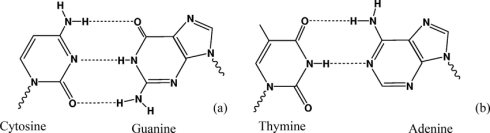
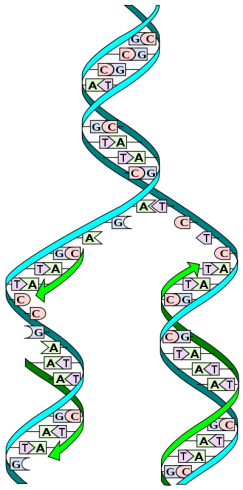
RNA و DNA ساختمانی نردبانی شکل دارند به این ترتیب که اسکلت این نردبان را واحدهای قند و فسفات به طور یک در میان با پیوندهای فسفودی استری 3 پریم و 5 پریم تشکیل داده و پله های آن را جفت بازهای آلی که از طریق پیوند هیدروژنی به هم متصل شده اند تشکیل می دهند. جفت باز ها بر اساس قوانین واتسون و کریک به هم اتصال می یابند بنابراین در مقابل هر باز پورین یک باز پیریمیدین قرار می گیرد. A مقابل T و C در مقابل G.
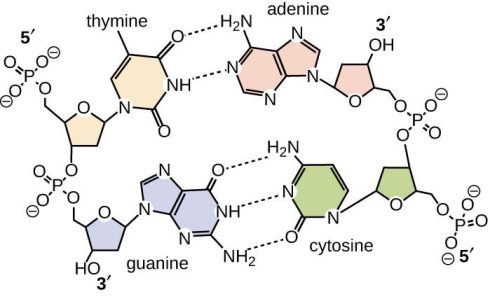
جفت بازهای CG از طریق سه پیوند و جفت باز های AT از طریق دو پیوند هیدروژنی کنار هم قرار می گیرند. با این شرایط تعداد T با تعداد A و تعداد C با تعداد G برابر می شود بنابراین می توان ترکیب باز های DNA را با ذکر درصد (GC درصدG + درصدC) در ترکیب آن مشخص نمود. برای مثال ترکیب DNA با ۵۶ GC٪ شامل ۲۸ G٪ ؛ ۲۸ C٪ ؛ ۲۲ T٪ و ۲۲ A٪ است.
ساختمان نردبانی شکل DNA حول یک محور فرضی چرخیده و ساختار مارپیچ شکل به خود می گیرد. این چرخش به همراه تابیدگی ملایم دو شیار کوچک و بزرگ را در DNA تشکیل می دهد.
به دلیل آنکه پیوند های فسفودی استری بین کربن 3 پریم از یک نوکلئوتید با فسفات کربن 5 پریم از نوکلئوتید مجاور تشکیل می شوند دو انتهای یک رشته DNA خطی با هم تفاوت دارند به این نحو که در یک انتها فسفات متصل به کربن 5 پریم آزاد بوده و در اتصال با نوکلئوتید دیگری قرار ندارد از طرف دیگر درانتهای 3 پریم نوکلئوتید دارای یک گروه 3 پریم OH آزاد خواهد بود که در تشکیل پیوند شرکت نکرده است.
بر اساس قوانین واتسون و کریک دو رشته DNA قرار گرفته روبروی هم نسبت به هم جهت عکس دارند و به فرم موازی ناهم سو هستند. دو رشته یک مولکول DNA دارای توالی های مکمل بوده به گونه ای که با دانستن توالی های بازی یک رشته می توان به آسانی توالی رشته مکمل را حدس زد. به طور معمول DNA را با نوشتن توالی بازی تنها یکی از رشته ها در جهت 3 → 5 که جهت سنتز RNA و یا DNA جدید از روی الگوی DNA می باشد توصیف می کنند.
چرخه سلولی
مراحلی که یک سلول از پایان یک تقسیم تا انتهای تقسیم بعدی می گذاراند را چرخه ی سلولی می نامند که شامل مراحل G1،G2،S و تقسیم (میتوز یا میوز) می باشد.G1 ، G2 و S مرحله ی استراحت یا اینترفاز نامیده می شوند. همانند سازی DNA یا مضاعف سازی DNA با فشردگی کم و ایجاد دو کروماتید خواهری در مرحله ی S اتفاق می افتد.

فشرده سازی DNA تا کروموزوم
تمام ژنوم هسته ای در چندین مولکول DNA خطی به نام کروموزوم جای گرفته است. اگر کل DNA یک سلول دیپلوئید انسان به طور کامل از هم باز شود طولی به اندازه ی 2 متر خواهد داشت!
این که این طول چگونه توانسته است در هسته سلولی به آن کوچکی قرار گیرد مدیون سیستم بسیار سازمان یابی شده ای برای بسته بندی DNA در قالب کروموزوم ها می باشد.

اولین سطح بسته بندی ساختاری به نام نوکلئوزوم است که در آن مولکول DNA دو دور اطراف 8 مولکول پروتئینی به نام هیستون می پیچد، هشت مولکول پروتئینی خود شامل دو مولکول از هر یک از هیستون های H2A ، H2B ، H3 و H4 که یک هسته ی اکتامری بشکه مانند را تشکیل می دهند، می باشد.
طول DNA پیچیده شده به دور هیستون حدود140 تا 150 جفت باز است و طول رشته ی DNA ی بین دو نوکلئوزوم مجاور بین 50 تا 70 جفت باز است که در مجموع تکرارهایی به طول تقریبا 190 تا 220 جفت باز تشکیل می شود.علاوه بر هیستون هایی که در اکتامر بشکه مانند قرار دارند،گروه دیگری از هیستون ها به نام هیستون های اتصالی وجود دارند (H1/H5 )که هرکدام از آنها به یک نوکلئوزوم متصل شده و کروماتوزوم را تشکیل می دهند. هیستون های اتصالی مانند یک گیره از باز شدن DNA پیچیده شده به دور نوکلئوزوم ها جلوگیری می کنند.
سطح بعدی فشرده سازی که شامل در هم تنیدگی واحدهای نوکلئوزوی می باشد رشته های پیچیده تری از DNA با قطری حدود 30 نانومتر می سازند. دو مدل برای فیبرهای 30 نانومتری پیشنهاد شده اند، یک مدل سیم پیچی(Solenoid model) و دو مدل روبان مارپیچ (Helical ribbon). فیبرهای 30 نانومتری شکل اصلی کروماتین هسته ای در مرحله ی اینترفاز می باشند.


در زمان تقسیم سلول ها، DNA به شکل بسیار متراکم تر و پیچیده تری در می آید که کروموزوم متافازی نام دارد، این کروموزوم ها را می توان با میکروسکوپ نوری مشاهده کرد. در چرخه ی سلولی پس از همانندسازی DNA ، کروموزوم های متافازی به وجود می آیند که هرکدام حاوی دو مولکول DNA و دو بازو به نام کروماتید می باشند که در محل سانترومر به هم متصل هستند.
در کروماتین (کمپلکس DNA-هیستون) DNA در برابر آنزیم های نوکلئاز محافظت می شود. نوکلئوزوم ها به صورت ساختارهای بسیار منظمی بسته بندی شده اند که نقش اساسی و مهمی در تعیین عملکرد ژن ها دارند.به طور کلی، دو نوع کروماتین اصلی وجود دارد:هتروکروماتین شکل غیرفعال که در آن نوکلئوزوم ها به صورت فشرده بسته بندی شده اند و یوکروماتین شکل فعال که ژن ها در آن جای دارند و به صورت شل بسته بندی شده اند. نواحی هتروکروماتین دارای توالی های بسیار تکرارشونده هستند و در مقایسه با یوکروماتین ژن های کمتری دارند.
میتوز و میوز:
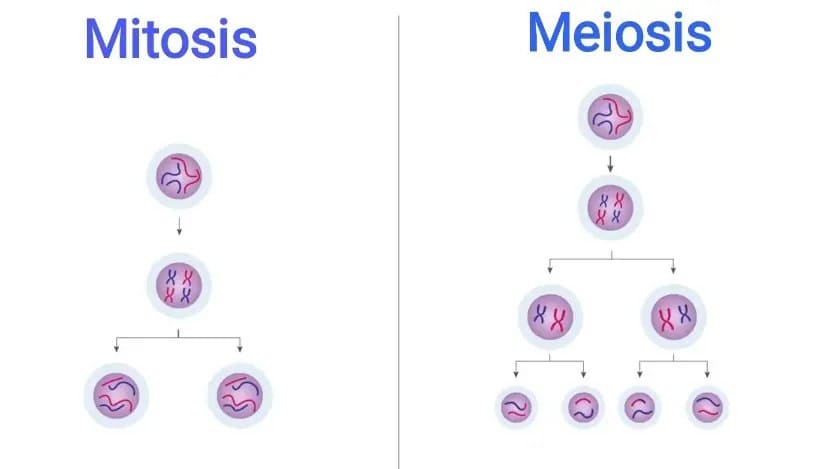
قبل از توضیح میتوز و میوز، ابتدا باید با مفاهیم هاپلوئید و دیپلوئیدآشنا شویم. در هسته ی سلول یا موجود هاپلوئید یک مجموعه ی کروموزومی (1n) وجود دارد در صورتی که موجود یا سلول دیپلوئید دو مجموعه ی کروموزومی (2n) در هسته ی خود دارد. همانطور که می دانید انسان یک موجود دیپلوئید است که دو مجموعه ی 23تایی، مجموعا 46 عدد کروموزوم در هسته ی خود دارد، اما گامت های انسان (تخمک و اسپرم) سلول های هاپلوئید با 23 عدد کروموزوم هستند که طی تقسیم میوز تولید می شوند.
به طور کلی موجودات زنده از طریق تقسیمات سلولی رشد کرده و تولید مثل می کنند.دو نوع تقسیم سلولی به نام های میتوز و میوز در بدن اتفاق می افتد. میتوز فرایندی است که طی آن سلول های جدید بدن(سوماتیک) ایجاد می شوند. هدف اصلی تقسیم میتوز رشد و جایگزین کردن سلولهای جدید با سلول های فرسوده است.
در حقیقت، تقسیم میتوز برای جایگزینی سلول هایی که طول عمر کوتاهی داشته و یا سلول هایی که دچار آسیب یا مرگ سلولی شده اند یک فرایند ضروری است و به موقع رخ ندادن میتوز می تواند منجر به تغییر در محتوای ژنومی سلول ها و اختلالات ژنتیکی شود. در میتوز یک سلول به دو سلول دختری دیپلوئید کاملا یکسان (Identical daughter cells) تقسیم می شود.
هدف از تقسیم میوز، تولید گامت ها یا سلول های جنسی اسپرم و تخمک است. در طول تقسیم میوز، یک سلول دو بار تقسیم شده و چهار سلول دختری هاپلوئید(23 کروموزومی) تولید می کند. بر خلاف تقسیم میتوز که در آن دو سلول با محتوای ژنتیکی کاملا یکسان تولید می شوند، در تقسیم میوز نوترکیبی هایی رخ می دهد که در نتیجه ی آن چهار سلول دختری که از نظر محتوای ژنتیکی متنوع هستند، ایجاد می شوند. اگر تقسیم میوز سلولهای دختری را دیپلوئید و بدون کاهش تعداد کروموزوم ها تولید می کرد، تعداد کروموزوم ها در نسل بعد دو برابر می شد.
تقسیم میتوز به طور کلی دارای چهار مرحله ی اصلی است:
1)پروفاز
2)متافاز
3)آنافاز
4)تلوفاز
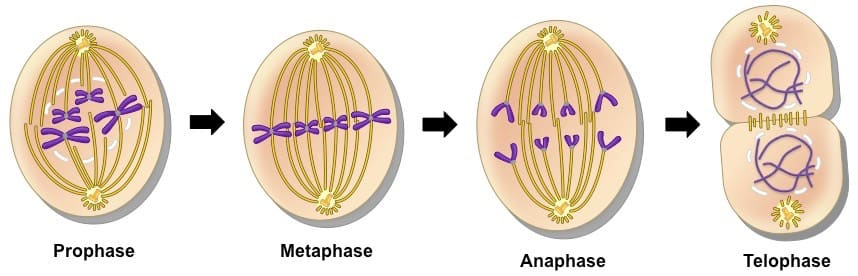
1)پروفاز:
در مرحله ی پروفاز، DNA به صورت بسته های بسیار محکم متراکم توسط پروتئین های هیستونی ساخته شده و قابل انتقال است. سانتریول های آشکار شده در سلول میکروتوبول ها را سازماندهی کرده و میکروتوبول ها به کروموزوم های DNA متصل می شوند.در مرحله ی پروفاز، غشای هسته که مولکول های DNA را از سیتوپلاسم جدا کرده بود، شکسته شده و میکروتوبول ها در ناحیه ی سانترومر کروموزوم ها به پروتئین های کینه توکور اتصال می یابند.
2)متافاز:
در متافاز،کروموزوم ها به وسیله ی میکروتوبول ها یی به نام دوک با نیروی برابر کشیده می شوند، و در مرکز سلول و منطقه ای به نام صفحه ی متافازی قرار می گیرند.
3)آنافاز:
در مرحله ی آنافاز میکروتوبول های دوک دوکروماتید خواهری یک کروموزوم را از هم جدا کرده و به سمت قطب های سلول می کشند.
4)تلوفاز:
زمانی که کروماتید های خواهری به وسیله ی رشته های دوک به سمت قطب ها ی سلولی کشیده شدند و به طور کامل به قطب های مخالف رسیدند، در سلول شکاف ایجاد شده و در مرحله ی تلوفاز تقسیم میتوز پایان می یابد. در ادامه ی تلوفاز سیتوکینز اتفاق افتاده و سیتوپلاسم هم بین دو سلول دختری تقسیم می شود.
فرایند میوز از دو تقسیم تشکیل شده است، میوز I ومیوز II ،که هر دوی این تقسیم ها چهار مرحله ی پروفاز،متافاز،آنافاز و تلوفاز تقسیم میتوز را دارا هستند.
میوزI
میوز I یک تقسیم کاهشی است که در آن کروموزوم های همولوگ از هم جدا می شوند.
پروفاز میوز I :کروموزوم ها فشرده شده، غشای هسته از بین رفته ، کروموزوم ها درکنار هم جفت شده و تترادها را تشکیل می دهند،در این مرحله بین کروموزوم های هومولوگ کراسینگ اور رخ داده و نوترکیبی ایجاد می شود.
پروفاز میوز I خود شامل 5 زیر مرحله است:
1)لیپتوتن
2)زیگوتن
3)پاکی تن
4)دیپلوتن
5)دیاکینز
لیپتوتن: اتفاقات این زیرمرحله شامل:افزایش حجم هسته ای،ظهور کروموزوم ها به صورت کشیده، تاب خورده و نازک.
زیگوتن: در این زیر مرحله کروموزوم های همولوگ جفت شده و سیناپس تشکیل می شود، تترادها (بی والانت ها) ی کروموزوم های همولوگ تشکیل می شود.
پاکی تن: مهم ترین اتفاق در زیرمرحله ی پاکی تن تشکیل کیاسماتا، کراسینگ اور و تبادل قطعات DNA بین دوکروماتید از کروموزوم های هومولوگ است.همچنین هستک از نظر اندازه رشد می کند.
دیپلوتن: در این زیرمرحله جدا شدن کروموزوم ها از یکدیگر آغاز شده، البته که در بعضی نقاط تبادل همچنان ادامه داشته و کروموزوم ها در این نقاط متصل باقی می مانند.
دیاکینز: در این زیرمرحله، کیاسما ناپدید شده، هستک و غشای هسته ناپدید شده و دوک ها تشکیل می شوند.
متافاز میوزI: کروموزوم های همولوگ در صفحه ی متافازی قرارگرفته و رشته های دوک به سانترومر کروموزوم ها متصل می شوند.
آنافاز میوزI: کروموزوم های همولوگ از همدیگر جدا شده و به سمت قطب های سلول می روند،در این مرحله بر خلاف آنافاز میتوز که در آن کروماتیدهای خواهری از هم جدا شده و هرکدام به سمت یک قطب سلول می روند، در آنافاز میوزI کروماتیدهای خواهری از هم جدا نمی شوند.
تلوفاز میوزI: غشای هسته در اطراف هر گروه کروموزوم ها تشکیل شده و دو هسته ی دختری قبل از ورود به دومین تقسیم میوز، برای مدتی در مرحله ی اینترفاز باقی می مانند.
مراحل میوزII به طور کامل مشابه با مراحل میتوز است، تنها تفاوتی که وجود دارد، تعداد کروموزوم هاست، تعداد کروموزوم ها در میتوز 46 عدد است،در حالی که تعداد کروموزوم ها در میوزII به 23 عدد رسیده است زیرا میوز l که یک تقسیم کاهشی بوده را پشت سر گذاشته اند.
فرایند همانندسازی:
همانندسازی به طور کلی از سه مرحله ی اصلی تشکیل شده است:
- باز شدن دو رشته DNA
- ساخت پرایمر و اتصال پرایمر به رشته ی الگو
- پلیمریزاسیون رشته DNA دختری
همانندسازی فرایندی است که از طریق آن یک مولکول دو رشته ای DNA مضاعف شده و از آن دو مولکول DNA دو رشته ای حاصل می شود. دانستن کامل همانندسازی و شیوه انجام آن در بدن به فهم دیگر روش های تکثیری DNA در محیط آزمایشگاه همچون PCR کمک شایانی خواهد کرد.
برای شروع ساخت DNA جدید ابتدا می بایست مبدا های همانند سازی خاصی شناسایی شود سپس پیوند های هیدروژنی دو رشته DNA در این محل گسسته شده و دو رشته از هم جدا شوند این عمل به وسیله آنزیم هلیکاز در سلول رخ می دهد و چنگال همانندسازی به شکل Y تشکیل می شود.
سپس هر رشته به عنوان الگو برای ساخت رشته جدید قرار خواهد گرفت در نتیجه دو رشته ای جدید تشکیل شده از یک رشته مادری و یک رشته دختری تازه سنتز شده به این نحو از همانندسازی فرم نیمه حفاظتی اطالق می شود. آنزیم DNA پلیمراز جهت ساخت رشته مکمل از چهار داکسی نوکلئوزید تری فسفات استفاده می کند. (dATP;dCTP;dGTP,dTTP).
آنزیم DNA polymerase برخالف آنزیم polymerase RNA به انتهای 3 پریم OH آزاد در ناحیه شروع همانندسازی برای ادامه رشته نیاز دارد و قادر به شروع ساخت رشته جدید از یک الگوی تک رشته ای برهنه نمی باشد. سر 3 پریم OH آزاد به وسیله یک قطعه کوتاه الیگونوکلئوتیدی به نام آغازگر یا پرایمر تامین می شود.
پرایمر از جنس RNA به وسیله آنزیم پرایماز ساخته می شود. رشته DNA در جهت 3 به 5 رشته رهبر (Leading)و رشته مقابل در جهت 5 به 3 رشته پیرو (Lagging)است.
جهت همانندسازی و سنتز رشته جدید همیشه از 5 پریم به 3 پریم است بنابراین جهت سنتز رشته رهبر در جهت حرکت چنگال همانندسازی است در حالیکه رشته پیرو در خلاف حرکت آنزیم پلیمراز قرار دارد بنابراین منقطع و ناپیوسته ساخته می شود. قطعات حاصل از همانندسازی رشته پیرو در نهایت بعد از حذف پرایمرهای RNA از ابتدای DNA های تازه سنتز شده به وسیله آنزیم لیگاز به یک دیگر متصل می شوند. از این رو همانندسازی را یک فرایند نیمه منقطع یا Discontinuous-Semi می نامند.
DNA به عنوان ماده کلیدی در ذخیره سازی و انتقال اطلاعات ژنتیکی و حمایت از عملکرد کروموزوم و داشتن رمزهای ارزشمند در غالب ژن برای ساخت RNA و پلی پپتیدهایی که بین سلول ها تفاوت ایجاد می کنند بسیار مطرح و قابل توجه هستند.
ژن ها قطعات DNA پراکنده ای هستند که در فاصله های متغیر در طول توالی DNA قرار دارند و به عنوان الگویی برای ساخت انواع مولکول های RNA مکمل مورد استفاده قرار می گیرند به فرایند ساخت RNA از روی DNA رونویسی و یا بیان ژن اطالق می شود.
پس از آن رونوشت اولیه RNA طی مراحلی بالغ شده و در نهایت یک RNA دارای عملکرد از روی رشته DNA ساخته خواهد شد. اساسا ترکیب توالی DNA در تمام سلول های یک جاندار یکسان است و در واقع تفاوت بین سلول ها به دلیل تفاوت در سطح بیان ژن ها در مرحله رونویسی است.
در این میان یک دسته از ژن ها بیان تقریبا ثابتی را در همه سلول های بدن نشان می دهند این ژن ها را به اصطلاح ژن های خانه دار یا Housekeeping می نامند ،در واقع عملکرد ژن های خانه دار برای حیات سلول ها ضروری و بنابراین بیان آنها در هر سلول اجباری است. برای مثال این ژن ها در فرایندهای پایه ای سلول همچون گلیکولیز و اسکلت سلولی نقش دارند. ژن های دیگر بیان اختصاصی بافتی نشان می دهند و یا اینکه در زمان های خاصی از مراحل تکوین و یا چرخه سلولی بیان می شوند.
فرایند رونویسی:
فرایندی که طی آن از یک دو رشته ای DNA یک تک رشته RNA ایجاد می شود را رونویسی و یا Transcription می نامند. هر دو رشته تشکیل دهنده DNA می توانند به عنوان الگو برای ساخت RNA قرار بگیرند.
رشته الگو را Template یا آنتی سنس Antisense و رشته غیر الگو را Nontemplate یا سنس Sense می نامند.
توالی RNA ی سنتز شده کاملا مشابه با رشته ی sense می باشد.(به استثنای تفاوت T و U). رونویسی برای انجام نیاز به توالی های تنظیمی مهمی در بالادست ژن دارد این توالی ها که تحت عنوان پروموتر نامیده می شوند محل شناسایی فاکتورهای رونویسی بوده که هدایت و فعالسازی آنزیم RNApolymerase را برعهده دارند. موجودات یوکاریوتی دارای سه گروه polymerase RNA هستند.
RNA POL I کدکننده ی RNA های ریبوزومی:
- 28 sr RNA
- sr RNA 18
- و sr RNA 5.8
RNA POL III کدکننده ی:
- 5 srRNA
- tRNA
- U6 sn RNA
- و دیگر انواع RNA های کوچک غیر کد کننده می باشد.
ژن های miRNA، mRNA و اغلب snRNA ها و snoRNA ها نیز توسط RNApol II کد می شوند.
- snRNA: RNA های کوچک هسته ای
- snoRNA: RNA های کوچک هستکی
- tRNA: RNA ناقل
- rRNA: RNA ریبوزومی
- mRNA: RNA پیک
پیرایش و پردازش RNA:
اکثر ژن های مهره داران (تقریبا تمام ژن های کدکننده پروتئین mRNA و برخی از ژن های RNA) اولین رونوشت RNA ای که از آن ها در سلول ساخته می شود خام بوده و باید تحت فرایندهای مشخصی بالغ شود. این عمل ها تحت عنوان پردازش (End modification) و پیرایش (Splicing) بیان می شوند.
RNA های ساخته شده توسط RNApol II دستخوش اصلاحاتی در انتهای 5 پریم و 3 پریم توالی قرار می گیرند. مدت کوتاهی بعد از شروع ساخت RNA اولیه که در ادامه به mRNA تبدیل می شود یک نوکلئوتید گوانین متیله، به جای پیوند معمول 3OH به 5P از طریق پیوند غیر طبیعی 5P به 5P به نوکلئوتید اول از سر 5 اتصال می یابد. به این فرایند کلاهک گذاری گفته می شود.
از جمله مهمترین نقش های این کلاهک می توان به محافظت انتهای 5 در برابر حمله نوکلئازها ، تسهیل پیرایش RNA و تسهیل انتقال mRNA از هسته به سیتوپلاسم را نام برد. سنتز محصولات آنزیم های RNApol I و RNApol III پس از شناسایی محل اختتام رونویسی متوقف می شود در حالی که انتهای مولکول های mRNA حاصل از RNApol II بعد از خاتمه رونویسی برش خورده و یک توالی شامل نوکلئوتیدهای آدنین پشت سر هم به انتهای 3 آن اضافه می شود.
طی فرایند پلی آدنیلاسیون حدودا 200 نوکلئوتید آدنین (AMP) به 3 اضافه شده و دم پلی A تشکیل می دهد. این دنباله پلی آدنینی در انتقال mRNAبه سیتوپلاسم و پایداری آن ،همینطور شناسایی mRNA توسط ماشین پروتئین سازی (ریبوزوم) کمک می کند.
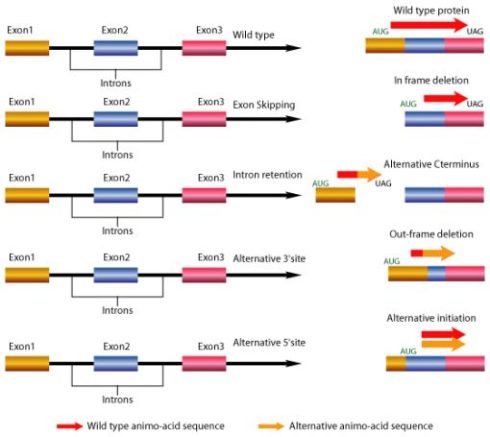
mRNA های اصلاح شده سپس باید در معرض پیرایش (Splicing)قرار گیرند. اسپلایسینگ فرآیندی است که طی آن اینترون ها حذف و اگزون ها به هم اتصال می یابند. اگزون ها دارای کدون های رمز کننده پروتئین هستند در حالی که هیچ رمز ژنتیکی کدکننده محصول نهایی در اینترون ها وجود ندارد.
اسپلایسینگ با شناسایی نواحی مرزی بین اگزون ها و اینترون ها آغاز می شود به نحوی که هر اینترون با یک دی نوکلئوتید GT آغاز شده و به یک دی نوکلئوتید AG ختم می شود. قانون (GT-AG) این نواحی Site Splice نامیده می شوند.
اسپلایس سایت ها به شدت حفاظت شده هستند. علاوه بر نواحی GT-AG راهنماهای دیگری نیز در اینترون وجود دارند تا به وسیله آنها فاکتورهای اسپلایسینگ در برش اینترون ها و اتصال صحیح اگزون ها دچار خطا نشوند. یکی دیگر از این توالی های راهنما تحت عنوان ناحیه انشعاب (Branch site) در فاصله حدودا 40 نوکلئوتیدی بالادست ناحیه AG واقع می باشد.
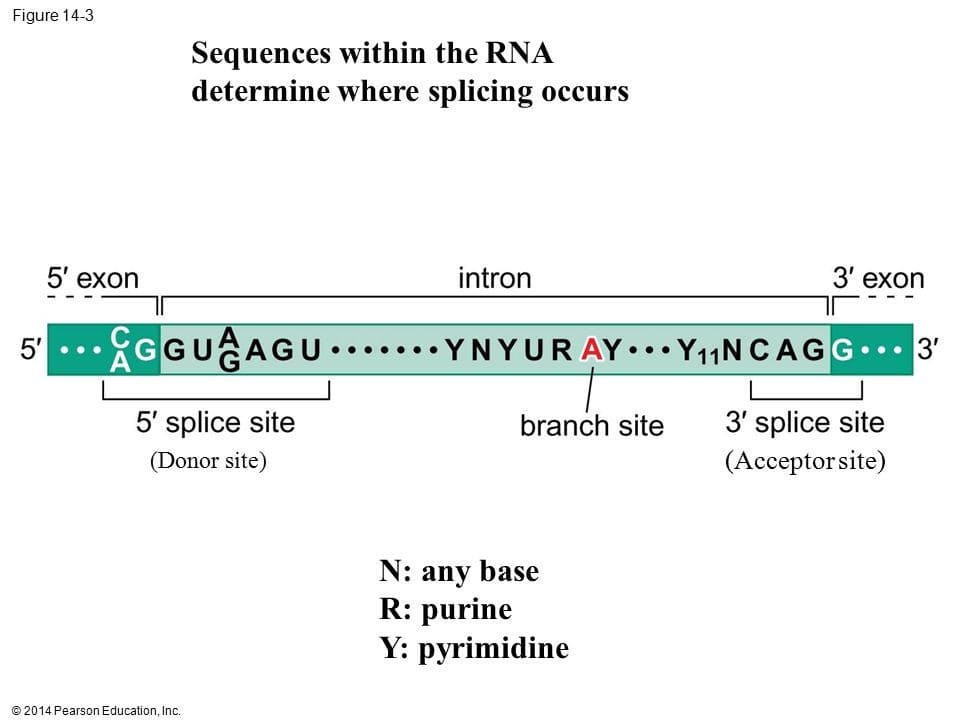
اسپلایسینگ به وسیله یک کمپلکس بزرگ پروتئینی به نام اسپلایسئوزوم انجام می شود.کمپلکس اسپلایسئوزوم دارای پنج نوع snRNA (U1,U2,U4,U5,U6) و بیش از 50 پروتئین است. مولکول های snRNA به همراه این پروتئین ها ذرات ریبونکلئوپروتئینی کوچک هسته ای (snurp or snRNP) را تشکیل می دهند.
توالی قسمتی از U1snRNA مکمل توالی مورد توافق جایگاه دهنده پیرایشی (donor site) می باشد در نتیجه این دو RNA به هم متصل می شوند از طرفی مولکول U2snRNA با جایگاه انشعاب پیوند می دهد سپس یک حمله نوکلئوفیلی نوکلئوتید G انتهای 5 را به نوکلئوتید A جایگاه انشعاب وصل کرده و یک ساختار کمند شکل را ایجاد می کند. حمله نوکلئوفیلی انتهای انتهای 3 اگزون بالادستی سبب برش و ازاد شدن RNA اینترونی به شکل یک کمند شده و دو قطعه RNA اگزونی به یکدیگر وصل می شوند.
Reference
- https://medpip.com/mag/6-%D8%AA%D9%81%D8%A7%D9%88%D8%AA-%D8%A8%DB%8C%D9%86-%D8%AA%D9%82%D8%B3%DB%8C%D9%85-%D9%85%DB%8C%D8%AA%D9%88%D8%B2-%D9%88-%D9%85%DB%8C%D9%88%D8%B2-%D8%AF%D9%88-%D9%81%D8%B1%D8%A7%DB%8C%D9%86%D8%AF/
- https://article.tebyan.net/265634/%D9%85%DB%8C%D9%88%D8%B2-1
- کتاب استراخان مولکولی 2
- کتاب ژنوم 3
- https://fa.wikipedia.org/wiki/%DA%86%D8%B1%D8%AE%D9%87_%D8%B3%D9%84%D9%88%D9%84
هزینه گرفتن مدرک ITE لندن برای دورهی ژنتیک مولکولی چقدر هست؟
سلام و وقت بخیر
جهت کسب اطلاعات و توضیحات تکمیلی با ازمایشگاه تماس بگیرید
برای دوره کاراموزی ژنتیک مولکولی . هزینشو لطف میکنید چون دوتا مبلغ هست و اینکه تاریخ شروعش کی میشه و تا چه زمانی ادامه داره
سلام و وقت بخیر
اطلاعات برگزاری دوره در همین صفحه ذکر شده است
جهت دریافت اطلاعات بیشتر با آزمایشگاه تماس بگیرید
شروع تا پایان دوره چقدر طول میکشد
سلام و وقت بخیر
دوره ژنتیک مولکولی 1ماه و نیم تا 2 ماه طول خواهد کشید