


اطلاعات عمومی,ویکی ژن
سیتوزول یا ماتریکس سیتوپلاسمی چیست؟
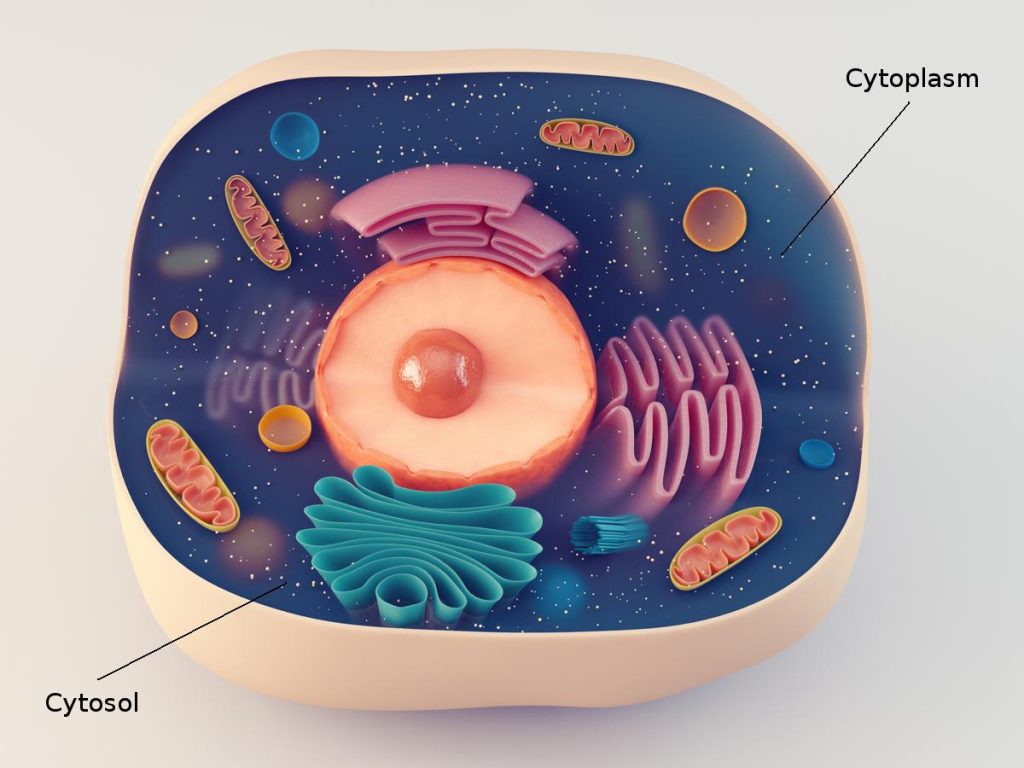
مقدمهای بر سیتوزول یا ماتریکس سیتوپلاسمی
- بسیاری از مولکولهایی که سلولها، بافتها و اندامها را تشکیل میدهند، با مولکولهای موجود در مواد غیرزنده یکسان هستند، در حالی که برخی دیگر منحصر به موجودات زندهاند.
- بیوشیمی (Biochemistry) مطالعه مولکولهای شیمیایی موجود در سیستمهای زیستی و واکنشهایی است که در آنها شرکت میکنند.
- بیولوژی مولکولی (Molecular biology) مطالعه ساختار و رفتار مولکولهای منفرد است. اگر “راز زندگی” در جای دیگری وجود داشته باشد، در درون همین مولکولها نهفته است.
- در واقع، همه سیستمهای زنده و غیرزنده تابع اصول فیزیکی و شیمیایی یکسانی هستند.
- درون سلولهای هر موجود زنده، ماده زنده یا پروتوپلاسم (Protoplasm) از مجموعهای از اجزای غیرزنده تشکیل شده است، از جمله پروتئینها، اسیدهای نوکلئیک، چربیها (لیپیدها)، کربوهیدراتها، ویتامینها، مواد معدنی، متابولیتهای زائد، تجمعهای بلورین و رنگدانهها که همگی از مولکولها و اتمهای تشکیلدهنده آنها ساخته شدهاند.
- پروتوپلاسم به دلیل آرایش و برهمکنش فوقالعاده پیچیده اجزای مختلف غیرزنده، زنده است. این شبیه به یک ساعت است که تنها زمانی به عنوان یک وسیله اندازهگیری زمان عمل میکند که همه چرخدندهها، فنرها و یاتاقانهای آن به شیوه خاصی چیده شده و با هم تعامل داشته باشند.
- با توجه به اصول فیزیکی جهانی، نه چرخدندههای ساعت و نه مولکولهای پروتوپلاسم نمیتوانند به گونهای با هم تعامل داشته باشند که این قوانین را نقض کند.
- هرچه درک ما از عملکرد پروتوپلاسم و عناصر آن بر اساس اصول شیمیایی بیشتر باشد، درک ما از پدیدههای زندگی نیز بیشتر خواهد شد.
- خارج از اندامکها، ماتریکس سیتوپلاسمی یا سیتوزول (Cytosol) جزء مایع و محلول سیتوپلاسم است. این فصل به بررسی ویژگیهای فیزیکی و شیمیایی سیتوزول اختصاص دارد.
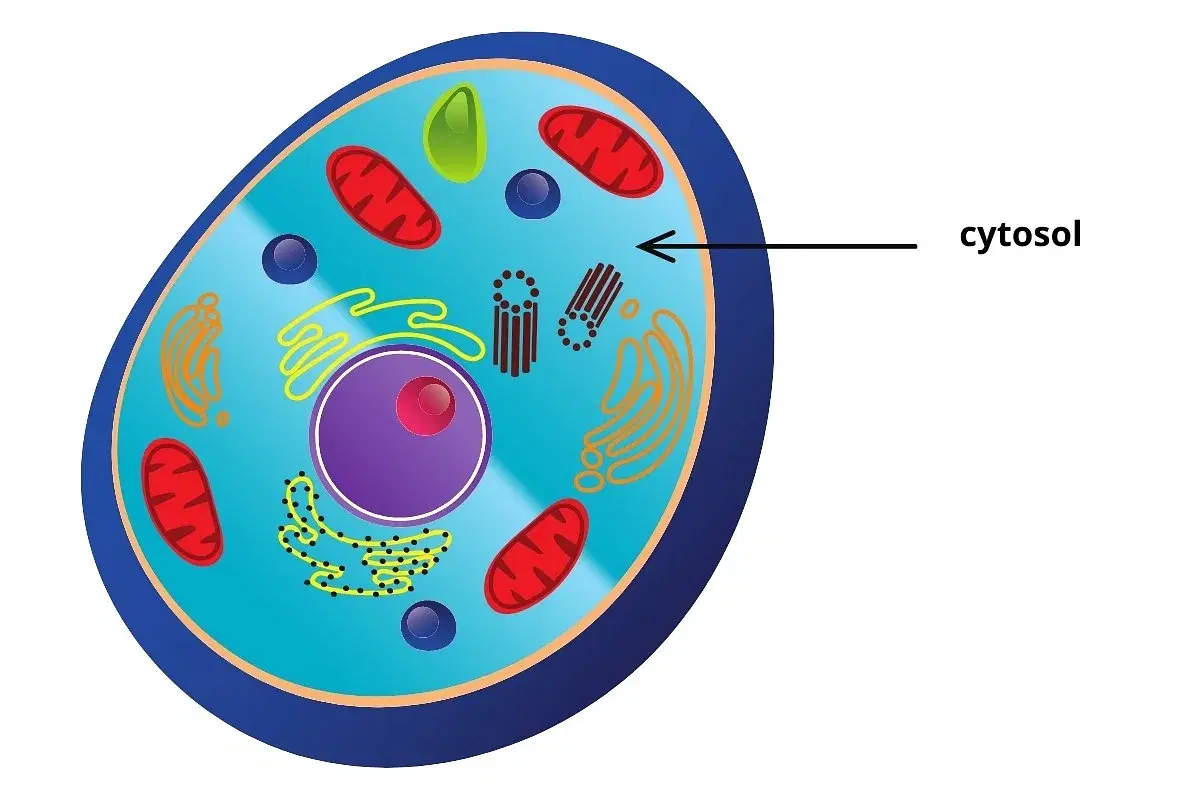
ماهیت فیزیکی سیتوزول (ماتریکس سیتوپلاسمی)
سیتوزول (ماتریکس سیتوپلاسمی) یک ماده کلوئیدی است که بیرنگ یا خاکستری، شفاف، چسبناک، ژلاتینی یا ژلهای شکل است. این ماده از آب متراکمتر بوده و میتواند جریان یابد. در گذشته بحثهای زیادی در مورد ماهیت فیزیکی ماتریکس وجود داشته است. در مورد خواص فیزیکی ماتریکس، پژوهشگران مختلف فرضیههای متفاوتی ارائه دادهاند. نظریههای آنها را میتوان به شکل زیر نشان داد:
- نظریه شبکهای (Reticular theory): طبق این نظریه، ماتریکس از شبکهای از الیاف یا ذرات در ماده زمینهای (Ground material) تشکیل شده است.
- نظریه حبابچهای (Alveolar theory): بوچلی در سال ۱۸۹۲ نظریه ح حبابچهای را مطرح کرد. طبق این نظریه، ماتریکس از چندین قطره معلق یا حبابچه (Alveoli) یا حبابهای ریز شبیه کف امولسیون تشکیل شده است.
- نظریه گرانولی (Granular theory): نظریه گرانولی اولین بار توسط آلتمان در سال ۱۸۹۳ ارائه شد. این فرضیه از این ایده حمایت میکند که ماتریکس از چندین گرانول (Granule) با اندازهها و پیکربندیهای مختلف تشکیل شده است. این ذرات به عنوان بیوپلاستها (Bioplasts) شناخته میشدند.
- نظریه رشتهای (Fibrillar theory): فلمینگ نظریه رشتهای را پیشنهاد کرد که ادعا میکند ماتریکس دارای ساختار رشتهای (Fibrillar) است.
تئوری کلوئیدی (Colloidal theory)
- پس از مطالعات میکروسکوپ الکترونی روی ماتریکس، به تازگی تئوری کلوئیدی مطرح شده است. طبق جدیدترین نظریه، ماتریکس بخشی محلول حقیقی و بخشی سیستم کلوئیدی است.
- محلول، مخلوطی از یک مایع به نام حلال و هر جزء شیمیایی جامد یا مایع به نام حل شونده است. ذرات حل شونده در یک محلول باید قطری کمتر از ۱/۱۰.۰۰۰ میلی متر داشته باشند. در بخش محلولی ماتریکس، مواد محلول مهم زیستی مانند گلوکز، اسیدهای آمینه، اسیدهای چرب، الکترولیتها، مواد معدنی، ویتامینها، هورمونها و آنزیمها در آب حل شدهاند.
- یک سیستم کلوئیدی به عنوان سیستمی توصیف می شود که حاوی یک محیط مایع است که ذرات با قطر تقریباً 1/1,000,000 to 1/10,000 millimetres در آن پراکنده شده اند. بنابراین، حالت کلوئیدی وضعیتی است که در آن یک ماده، مانند پروتئین یا مولکول بزرگ دیگر، در ماده دیگری پراکنده شده و چندین فاز ریز در یک فاز پیوسته واحد معلق می شود.
- هر سیستم کلوئیدی از دو فاز تشکیل شده است: فاز غیرپیوسته یا فاز پراکنده و فاز پیوسته یا فاز پراکنده کننده. کل پروتوپلاسم (سیتوپلاسم + هسته پلاسم) یک محلول کلوئیدی است زیرا اجزای اصلی مولکولی پروتوپلاسم، پروتئین ها، تمام خواص یک حالت کلوئیدی را نشان می دهند.
- پروتئینها به دلیل دو عامل، کلوئیدهای پایداری را تشکیل میدهند: اول اینکه آنها یونهای باردارشدهای هستند که در محلول همدیگر را دفع میکنند و دوم اینکه هر مولکول پروتئین مولکولهای آب را در لایههای متمایز در اطراف خود جذب میکند.
تغییر فاز (Phase Reversal)
- سیتوزول (ماتریکس سیتوپلاسمی)، مانند بسیاری از سیستمهای کلوئیدی، تغییر فاز را نشان میدهد. برای مثال، ذرات ژلاتین (فاز غیرپیوسته) در آب (فاز پیوسته) در یک قوام رقیق و لرزان پخش می شوند.
- این حالت به عنوان سول (Sol) شناخته می شود. با سرد شدن محلول، ژلاتین به فاز پیوسته تبدیل می شود و آب به فاز غیرپیوسته تبدیل می شود.
- علاوه بر این، محلول اکنون نیمه جامد شده و به عنوان ژل (Gel) شناخته می شود. در حالت ژل، مولکولهای یک ماده کلوئیدی توسط انواع پیوندهای شیمیایی از جمله برهمکنشهای هیدروژنی (H-H)، کربن-هیدروژن (C-H) و کربن-نیتروژن (C-N) به هم متصل می شوند.
- پیوندهای شیمیایی ماهیت و استحکام پایداری ژل را تعیین می کنند. گرم کردن محلول ژل آن را دوباره به مایع تبدیل کرده و فازها را معکوس می کند.
- در شرایط عادی، تغییر فاز سیتوزول (ماتریکس سیتوپلاسمی) به فرآیندهای فیزیولوژیکی، مکانیکی و بیوشیمیایی سلول بستگی دارد.
سازماندهی شیمیایی سیتوزول (ماتریکس سیتوپلاسمی)
ماتریکس سیتوپلاسمی از چندین جزء شیمیایی به صورت اتمها، یونها و مولکولها تشکیل شده است.
عناصر شیمیایی (Chemical Elements)
- تقریباً 46 مورد از 92 عنصر طبیعی موجود در سیتوزول (ماتریکس سیتوپلاسمی) یافت می شوند.
- بیست و چهار مورد از این عناصر برای زندگی ضروری تلقی می شوند (عناصر ضروری نامیده می شوند)، در حالی که برخی دیگر تنها به دلیل وجود در محیط اطراف ارگانیسم در سیتوزول وجود دارند.
- از میان بیست و چهار جزء اصلی، شش عنصر نقش های حیاتی در سیستم های بیولوژیکی ایفا می کنند. اینها عناصر اصلی هستند: کربن (20٪)، هیدروژن (10٪)، نیتروژن (3٪)، اکسیژن (62٪)، فسفر (1.14٪) و گوگرد (S، 0.14 درصد).
- اکثر مولکول های آلی از این شش جزء تشکیل شده اند. پنج عنصر مهم باقیمانده که به مقدار کمتر در سیستم های زنده یافت می شوند، کلسیم (Ca، 2.5٪)، پتاسیم (K، 0.11٪)، سدیم (Na، 0.10٪)، کلر (Cl، 0.16٪) و منیزیم (Mg، 0.16٪) هستند. (Mg، 0.07 درصد).
- عناصر دیگر، که به عنوان عناصر کمیاب شناخته می شوند، نیز به مقدار کم در حیوانات و گیاهان وجود دارند، اما برای ادامه حیات ضروری هستند.
- این عناصر شامل آهن (0.10٪)، ید (0.014٪)، مولیبدن (Mo)، منگنز (Mn)، کبالت (Co)، روی (Zn)، سلنیوم (Se)، مس (Cu)، کروم (Cr)، قلع (Sn)، وانادیم (V) می شود. ، سیلیسیم (Si)، نیکل (Ni)، فلوئور (F) و بور (B).
- ماتریکس سیتوپلاسمی از انواع مختلف یون ها تشکیل شده است. یون ها برای حفظ فشار اسمزی و تعادل اسید-باز در سلول ها ضروری هستند.
- نگهداری یون ها در ماتریکس منجر به افزایش فشار اسمزی و در نتیجه ورود آب به سلول می شود.
- مایع داخل سلولی (ماتریکس) حاوی غلظت متفاوتی از یون ها نسبت به مایع بین سلولی است. غلظت K+ و Mg++ می تواند داخل سلول بالا باشد، اما غلظت Na+ و Cl- می تواند خارج سلول بالا باشد.
- اختلاف زیادی بین K+ داخل سلولی و Na+ خارج سلولی در سلول های عضلانی و عصبی وجود دارد.
- ممکن است یون های آزاد کلسیم (Ca++) در سلول ها یا خون وجود داشته باشند. سلول های اپیتلیال علف ها حاوی یون های سیلیسیم هستند.
- یون های آزاد فسفات (اولیه، H2PO4، و ثانویه، HPO4) در ماتریکس و خون یافت می شوند. این یون ها به عنوان بافر عمل کرده و pH خون و مایعات سلولی را تثبیت می کنند.
- علاوه بر سولفات (SO4 —)، کربنات (CO3 —)، بیکربنات (HCO3 —)، منیزیم (Mg++ ) و اسیدهای آمینه، یون های سلول های مختلف نیز شامل کربنات (CO3 —)، بیکربنات (HCO3 —) و کربنات (CO3 —) می شوند.
الکترولیتها و غیرالکترولیتها (Electrolytes and Non-electrolytes)
ماتریکس از الکترولیتها و غیرالکترولیتها تشکیل شده است.
(۱) الکترولیتها (Electrolytes)
- الکترولیتها برای حفظ فشار اسمزی و تعادل اسید-باز در ماتریکس ضروری هستند.
- نمونههایی از الکترولیتها یونهای Mg2+، فسفات و سایر مواد هستند.
(۲) غیرالکترولیتها (Non-electrolytes)
- برخی از مواد معدنی در ماتریکس به صورت غیر یونی کننده وجود دارند.
- غیرالکترولیتهای ماتریکس شامل Na، K، Ca، Mg، Cu، I، Fe، Mn، Fl، Mo، Cl، Zn، Co و Ni و غیره هستند.
- آهن (Fe) در هموگلوبین، فریتین، سیتوکرومها و برخی آنزیمها از جمله کاتالاز و سیتوکروم اکسیداز وجود دارد. کلسیم (Ca) را میتوان در خون، ماتریکس و استخوانها یافت.
- مس (Cu)، منگنز (Mn)، مولیبدن (Mo) و روی (Zn) به عنوان کوفاکتور برای فرآیندهای آنزیمی عمل میکنند.
- ید و فلوئور به ترتیب برای متابولیسم تیروئید و مینای دندان ضروری هستند.
ترکیبات غیرآلی سیتوزول (Inorganic Compounds of Cytosol)
ترکیبات غیرآلی شامل عناصر، فلزات، غیرفلزات و ترکیبات آنها مانند آب، نمک ها و دامنه وسیعی از الکترولیت ها و غیرالکترولیت ها می شود. در بخش قبلی به طور گسترده به مواد غیرآلی به جز آب پرداخته شد که در پاراگراف بعدی به آن اشاره خواهد شد.
آب (Water)
آب فراوانترین جزء غیرآلی سیتوزول است (استثناهای قابل توجه، دانهها، استخوان و مینهی دندان هستند). آب بین ۶۵ تا ۸۰ درصد از ماتریکس را تشکیل میدهد. در ماتریکس دو شکل از آب وجود دارد: آب آزاد و آب متصل (bound water).
- آب آزاد: به ۹۵ درصد از کل آب سلولی اشاره دارد که توسط ماتریکس به عنوان حلال برای مولکولهای غیرآلی و آلی مختلف مورد استفاده قرار میگیرد.
- آب متصل (bound water): باقیمانده ۵ درصد از کل آب سلولی است که به طور ضعیفی با پیوندهای هیدروژنی یا عوامل دیگر به مولکولهای پروتئین متصل است.
میزان آب ماتریکس سلولی یک ارگانیسم به طور مستقیم به سن، محیط زیست و فعالیت متابولیک آن بستگی دارد. برای مثال، سلولهای جنین حاوی ۹۰ تا ۹۵ درصد آب هستند، در حالی که سلولهای ارگانیسم بالغ به تدریج آب کمتری دارند. به طور کلی، سلولهای موجودات آبزی پایینتر نسبت به سلولهای جانوران خشکیزی بالاتر، حاوی نسبت بیشتری از آب هستند. علاوه بر این، نسبت آب در ماتریکس بسته به سرعت متابولیک از سلولی به سلول دیگر متفاوت است.
ساختار مولکولی آب (Molecular structure of water)
ویژگیهای فیزیکی منحصر به فرد آب، در ساختار مولکولی آن منعکس میشود. هیدروژن و اکسیژن از طریق تشکیل پیوندهای کووالانسی که در آن اتمها جفت الکترونهایی را برای تشکیل پیوندهای شیمیایی به اشتراک میگذارند، با هم ترکیب شده و آب را تشکیل میدهند.
پیوندهای کووالانسی، پیوندهای شیمیایی قوی بین اتمها هستند که حاوی مقدار قابل توجهی انرژی شیمیایی (۱۱0.6 کیلوکالری بر مول یا ۴۶۲ کیلوژول بر مول) هستند. هیدروژن در شکل با تک الکترون خود نشان داده شده است که میتواند آن را با یک اتم اکسیژن به اشتراک بگذارد. هر اتم اکسیژن میتواند دو الکترون را با دو اتم هیدروژن به اشتراک بگذارد.
ویژگی های فیزیکی منحصر به فرد آب و کاربرد بیولوژیکی آنها
برخی از ویژگی های استثنایی آب باعث می شود تا برای نقش حیاتی خود در سیستم های پروتوپلاسمی (یعنی سیتوزول یا ماتریکس) کاملا مناسب باشد. از جمله ویژگی های متمایز آب می توان به موارد زیر اشاره کرد:
آب به عنوان حلال (Water as a solvent)
- آب باثبات ترین و انعطاف پذیرترین حلال به شمار می رود. ماهیت قطبی آب، ویژگی های آن را به عنوان حلال برای مواد معدنی مانند یون های معدنی، جامدات و غیره و ترکیبات آلی مانند کربوهیدرات ها و پروتئین ها تعیین می کند.
- آب به دلیل قطبی بودن می تواند به طور الکترواستاتیکی به گروه های پروتئینی با بار مثبت و منفی متصل شود. بنابراین، هر گروه آمینی در یک مولکول پروتئین می تواند به 2.60 مولکول آب متصل شود. از آنجایی که تمام واکنشهای شیمیایی سلولی در محلول آبی رخ میدهند، بنابراین حلالیت اهمیت بیولوژیکی بسیار زیادی دارد.
- آب همچنین به عنوان یک محیط پراکنده موثر برای ساختار کلوئیدی ماتریکس عمل می کند.
ویژگی های حرارتی آب (Water’s thermal properties)
- آب تنها ماده ای است که در محدوده دمایی طبیعی زمین در سه حالت جامد، مایع و گاز وجود دارد. افزایش دمای 1 گرم آب به میزان 1 درجه سانتیگراد (مثلاً از 15 درجه سانتیگراد به 16 درجه سانتیگراد) به 1 کالری (4.185 ژول) انرژی نیاز دارد.
- چنین ظرفیت حرارتی بالایی از آب تأثیر تعدیل کننده قابل توجهی بر تغییرات دما در محیط دارد و یک عامل نجات دهنده عالی برای زندگی است. علاوه بر این، آب گرمای تبخیر بالایی دارد.
- برای تبدیل 1 گرم آب مایع به بخار آب به بیش از 540 کالری (2259 ژول) انرژی نیاز است. در نتیجه، دمای جوش آب (100 درجه سانتیگراد) برای ماده ای با جرم مولکولی نسبی پایین به طور شگفت انگیزی بالا است.
- به نظر می رسد اگر این اتفاق خوشایند رخ نمی داد، به احتمال زیاد آب مایع هرگز روی زمین وجود نداشت و به فضا فرستاده می شد. همچنین، برای گیاهان و جانوران خشکی زی، خنک شدن ناشی از تبخیر آب، یک تکنیک ضروری برای حذف گرمای اضافی است.
- علاوه بر این، در دمای بسیار پایین، مقادیر زیادی انرژی (335 ژول یا 80 کالری به ازای هر گرم) باید برای تبدیل آب از مایع به جامد هدر رود. این پدیده به عنوان گرمای ذوب شناخته می شود.
- دمای صفر درجه سانتیگراد دمای ذوب آب است. رفتار عجیب و غریب چگالی آب به عنوان تابعی از تغییر دما، ویژگی مهم دیگری از نظر زیست شناسی است.
- بیشتر مایعات با سرد شدن چگالی بیشتری پیدا می کنند. با این حال، آب در دمای 4 درجه سانتیگراد به حداکثر چگالی خود می رسد و سپس با کاهش دما سبک تر می شود.
- در نتیجه، یخ به جای اینکه در کف دریاچه ها و حوضچه ها فرو رود، شناور می شود. این امر از یخ زدن زندگی آبزی جلوگیری می کند.
کشش سطحی (Surface tension)
کشش سطحی آب زیاد است. این ویژگی که ناشی از چسبندگی بالای مولکول های آب است، برای حفظ شکل و حرکت پروتوپلاسمی ضروری است. ایسکوزیتگی پایین آب، با وجود کشش سطحی بالای آن، عبور خون از مویرگ های ریز و سیتوپلاسم درون مرزهای سلولی را تسهیل می کند. مولکولهای حل شده در آب تمایل دارند در مرز بین فاز مایع آن و سایر فازها جمع شوند و کشش سطحی آب را کاهش دهند. این ممکن است نقش کلیدی در تشکیل غشای پلاسمایی داشته باشد و نقش اساسی در انتقال مولکول ها از آن داشته باشد.
شفافیت (Transparency)
آب در برابر نور شفاف است و این امکان را برای اندامکهای تخصصی فتوسنتز، کلروپلاستها، درون سلول گیاهی برای جذب نور خورشید جهت فتوسنتز فراهم میکند.
ترکیبات آلی سیتوزول (Organic Compounds of Cytosol)
ترکیبات آلی، مواد حاوی کربن (C) همراه با یک یا چند عنصر اضافی مانند هیدروژن (H)، نیتروژن (N) یا گوگرد (S) هستند. ترکیبات آلی به طور معمول از مولکول های بزرگی تشکیل شده اند که توسط مونومرها با ساختار واحد مشابه یا متمایز ساخته می شوند. مونومر (یونانی: mono=یک؛ meros=بخش) کوچکترین واحد یک مولکول آلی است که ممکن است به تنهایی وجود داشته باشد.
برخی از مولکولهای آلی مانند کربوهیدراتها به عنوان مونومر در ماتریکس عمل میکنند. به طور معمول، مونومرها با مونومرهای دیگر برای تشکیل الیگومرها (یونانی: oligo=چند یا کوچک) و پلیمرها (یونانی: poly=بسیار) پیوند برقرار میکنند. الیگومرها از تعداد کمی مونومر تشکیل شده اند، در حالی که پلیمرها از تعداد زیادی مونومر تشکیل شده اند.
الیگومرها و پلیمرها از مولکول های بزرگ یا ماکرومولکول ها تشکیل شده اند. هنگامی که ماکرومولکول یک پلیمر از انواع یکسان مونومر تشکیل شده باشد، به آن هموپلیمر می گویند، اما زمانی که از انواع مختلف مونومر تشکیل شده باشد، به آن هتروپلیمر می گویند. کربوهیدراتها، لیپیدها، پروتئینها، ویتامینها، هورمونها و نوکلئوتیدها اجزای اصلی آلی ماتریکس را تشکیل میدهند.
1. کربوهیدراتها (Carbohydrates)
کربوهیدرات ها (لاتین، carbo=کربن یا زغال سنگ، یونانی، hydro=آب) مولکول هایی بر پایه کربن، هیدروژن و اکسیژن هستند. آنها منبع اصلی انرژی برای همه موجودات زنده هستند. تنها قسمتهای سبز گیاهان و تعداد کمی از میکروارگانیسمها قادرند از طریق فتوسنتز در حضور نور و کلروفیل، کربوهیدرات را از آب، دی اکسید کربن و کلروفیل تولید کنند.
تمام حیوانات، قسمت های غیرسبز گیاه (مثلاً ساقه، ریشه)، گیاهان غیرسبز (مثلاً قارچ)، باکتری ها و ویروس ها برای نیاز گلوکز خود به قسمت های سبز گیاه وابسته هستند. کربوهیدرات ها به لحاظ شیمیایی پلی هیدروکسی آلدهید یا کتون هستند و به شرح زیر دسته بندی می شوند: الف. مونوساکاریدها (مونومرها)، ب. الیگوساکاریدها (الیگومرها)، و ج. پلی ساکاریدها (پلیمرها).
الف. مونوساکاریدها (Monosaccharides)
مونوساکاریدها با فرمول تجربی Cn(H2O)n، قندهای ساده هستند. آنها بر اساس تعداد اتم های کربن در مولکول هایشان به شرح زیر طبقه بندی و نامگذاری می شوند:
- تریوزها: مولکولهایی با سه اتم کربن دارند، مانند گلیسראלدهید و دی هیدروکسی استون.
- تتروزها: مولکولهای حاوی چهار اتم کربن هستند، مانند اریترولوز و اریتروز.
- پنتوزها: مولکولهای آنها شامل ریبوز، ریبولوز، دئوکسی ریبوز، آرابینوز و زایلولوز است که پنج اتم کربن دارند.
- هگزوزها: گلوکز، مانوز، فروکتوز و گالاکتوز نمونههایی از هگزوزها با شش اتم کربن در مولکولهایشان هستند.
- هپتوزها: مانند سدوهپتولوز، حاوی هفت اتم کربن در مولکولهایشان هستند.
به طور معمول، مونوساکاریدها به صورت ایزومر وجود دارند. برای مثال، سه قند هگزوز – گلوکز، فروکتوز و گالاکتوز – حاوی تعداد مشابهی از اتمهای کربن، هیدروژن و اکسیژن (یعنی C6H12O6)هستند، اما به دلیل پیکربندیهای اتمی منحصر به فرد در مولکولهایشان، قندهای متمایزی محسوب میشوند. گلوکز و گالاکتوز به ترتیب ایزومرهای نوری و فضایی هستند.
اگر یک اتم کربن در مولکولی با چهار گروه شیمیایی مجزا که به آن متصل هستند وجود داشته باشد، این گروهها میتوانند به دو روش جایگزین در اطراف اتم کربن (چنین اتم کربنی اغلب اتم کربن نامتقارن نامیده میشود) به صورت فضایی مرتب شوند.
این دو پیکربندی متمایز به عنوان تصاویرآینه ای شناخته می شوند و یک تصویر مفید از چنین ساختارهایی دو دست انسان است که به طور یکسان شکل گرفته اند اما قابل روی هم قرار گرفتن نیستند. این دو ایزومر به طور مشابه با آلدوتریوزهای D و L گلیسرآلدهید به عنوان ‘D’ یا ‘L’ نشان داده میشوند.
اکثر مونوساکاریدها از نظر نوری فعال هستند، یعنی کربن (های) نامتقارن آنها باعث چرخش صفحه نور پلاریزه می شود. مولکولهایی که صفحه پلاریزاسیون را به سمت راست میچرخانند، همانطور که به سمت منبع نور نگاه میکنید، به عنوان دکستروراتوری شناخته میشوند و با نماد d یا (+) نشان داده میشوند، در حالی که وضعیت مخالف، لِوُراتوری، با نماد l یا (-) نشان داده میشود. (-) .
لازم به یادآوری است که D و L بزرگ به ساختار اشاره دارند، در حالی که d و l کوچک به فعالیت نوری اشاره دارند که قبل از تعیین ساختار ایجاد شده است. در نتیجه، به D (+) – گلوکز، که به عنوان دکستروز نیز شناخته می شود، و D (-) – فروکتوز، که به عنوان لولوز نیز شناخته می شود، اشاره می شود.
علاوه بر این، قندها را می توان برای سادگی به صورت یک زنجیره مستقیم خطی نشان داد. در واقعیت، پیکربندی حلقوی مرتبط تر است. این یک ایزومر با یک پل اکسیژن است که دو تا از کربن ها را به هم متصل می کند.
ایجاد حلقه در موقعیت یک، یک کربن نامتقارن جدید ایجاد می کند. به دلیل استریوشیمی مونوساکاریدها، حلقه ایجاد شده دارای پنج یا شش عضو است. یک حلقه با هفت عضو بیش از حد تحت فشار قرار می گیرد. در قندهای پنتوز (پنج کربنه) مانند ریبوز، یک حلقه فورانوز با پنج عضو ایجاد می شود. در هگزوزهایی مانند فروکتوز و گلوکز، یک حلقه پیرانوز با شش عضو ساخته می شود. هاورث روش مؤثری برای نشان دادن ساختارهای حلقوی قندها پیشنهاد کرد (1927).
بنابراین، در گلوکوپیرانوز، اتمهای کربن 2 و 3 جلوی صفحه کاغذ قرار دارند، در حالی که اتم کربن 5 و اکسیژن حلقه پشت صفحه کاغذ قرار دارند. گروههای جانشین یا بالای صفحه حلقه یا پایین آن قرار دارند.
مونوساکاریدها که مونومر هستند، قابل تقسیم یا هیدرولیز بیشتر به مولکولهای سادهتر نیستند. پنتوزها و هگزوزها فراوانترین مونوساکاریدهای موجود در ماتریکس هستند.
RNA و برخی از کوآنزیمها مانند نیکوتینامید آدنین دینوکلئوتید (NAD)، نیکوتینامید آدنین دینوکلئوتید فسفات (NADP)، آدنوزین تری فسفات (ATP) و کوآنزیم A (CoA) شامل ریبوز، یک قند پنتوز هستند.
دی اکسی ریبوز، یک قند پنتوز، جزء ضروری اسید دئوکسی ریبونوکلئیک (DNA) است.
ریبولوز یک قند پنتوز است که برای فرآیند فتوسنتز مورد نیاز است. گلوکز، یک قند هگزوز، منبع اصلی انرژی سلول است. فروکتوز و گالاکتوز دو قند هگزوز ضروری دیگر ماتریکس هستند.
ب. الیگوساکاریدها (Oligosaccharides)
مولکولهای الیگوساکاریدها شامل دو تا ده مونوساکارید (مونومر) هستند. مونومرها از طریق اتصالات یا پیوندهای گلیکوزیدی به هم متصل باقی میمانند. الیگوساکاریدهای مهم عبارتند از:
- دی ساکاریدها شامل دو مونومر، به عنوان مثال ساکارز، مالتوز، لاکتوز و غیره.
- تری ساکاریدها حاوی سه مونومر، مانند رافینوز، مانوتریوز، رابینوز، رامنوز، جنتیانوز و مליטوز هستند.
- تترا ساکاریدها مانند استاکیوز و اسکوردوز از چهار مونومر تشکیل شده اند.
- پنتا ساکاریدها شامل پنج مونومر، به عنوان مثال، ورباسکوز هستند.
دی ساکاریدهایی مانند ساکارز، مالتوز و لاکتوز رایج ترین الیگوساکاریدهای موجود در سلول های حیوانی و گیاهی هستند. ساکارز و مالتوز به طور عمده در ماتریکس سلول های گیاهی یافت می شوند، در حالی که لاکتوز فقط در ماتریکس سلول های حیوانی وجود دارد. مولکول های ساکارز از D-گلوکز و D-فروکتوز تشکیل شده اند. مولکولهای مالتوز از دو مولکول D-گلوکز تشکیل شدهاند. مولکولهای لاکتوز از دو مونومر یعنی D-گلوکز و D-گالاکتوز تشکیل شدهاند. مشابه مونوساکاریدها، همه دی ساکاریدها شیرین، محلول در آب و قابل تبلور هستند.
ج. پلی ساکاریدها (Polysaccharides)
پلی ساکاریدها در مولکولهای درشت خود از دهها تا هزاران مونوساکارید به عنوان مونومر تشکیل شدهاند. آنها فرمول تجربی (C6H10O6)n را دارند. مولکولهای پلی ساکارید از نظر اندازه کلوئیدی بوده و دارای وزن مولکولی بالایی هستند. هیدرولیز پلی ساکاریدها به قندهای ساده امکان پذیر است. دو نوع عملکردی اصلی برای پلی ساکاریدها وجود دارد: پلی ساکاریدهای ساختاری و پلی ساکاریدهای تغذیهای.
به طور عمده، پلی ساکاریدهای ساختاری به عنوان اجزای ساختاری خارج سلولی یا داخل سلولی عمل میکنند. سلولز (موجود در دیواره سلولی گیاهان)، مانان (یک هموپلیمر از مانوز که در دیواره سلولی مخمر یافت میشود)، کیتین (در اسکلت خارجی بندپایان و دیواره سلولی اکثر قارچها و برخی جلبکهای سبز)، اسید هیالورونیک، سولفات کراتین و سولفات کندرویتین (به ترتیب در غضروف و سایر بافتهای همبند یافت میشوند) و پپتیدوگلیکانها (در دیواره سلولی باکتری) در این دسته قرار دارند.
پلی ساکاریدها به عنوان ذخایر مونوساکارید عمل کرده و تحت گردش دائمی متابولیک قرار میگیرند. این دسته شامل نشاسته (سلولهای گیاهی و باکتریها)، گلیکوژن (سلولهای حیوانی)، inulin (کنگر فرنگی و قاصدک) و پارامیلوم (یک هموپلیمر ذخیرهای و غیر شاخهای از گلوکز که به عنوان منبع انرژی در برخی پروتیستها مانند یوجلنا یافت میشود) هستند.
برخی از مولکولهای پلی ساکارید زنجیرههای غیر شاخهای (یعنی خطی) با ساختارهای نواری یا مارپیچی (معمولاً مارپیچ چپگرد) هستند. برخی دیگر از پلی ساکاریدها مانند بسیاری از پروتئینها، شاخهای بوده و شکل کروی به خود میگیرند. بر اساس ساختار شیمیایی آنها، پلی ساکاریدها را میتوان به دو دسته اصلی طبقه بندی کرد: هموپلی ساکاریدها و هتروپلی ساکاریدها.
د. هموپلی ساکاریدها (Homopolysaccharides)
مولکولهای هموپلی ساکاریدها حاوی انواع مشابهی از مونوساکاریدها هستند. مهمترین هموپلی ساکاریدهای ماتریکس، نشاسته، گلیکوژن، پارامیلوم و سلولز هستند.
نشاسته (Starch)
- نشاسته یک پلیمر ذخیرهسازی سلول گیاهی تغذیهای است (به عنوان مثال، غدههای سیب زمینی). به طور معمول، به صورت دانه یا گرانول در سلولها ظاهر میشود (آنها در داخل پلاستیدهای کروی شکل قرار دارند). گرانولهای نشاسته حاوی مخلوطی از دو پلی ساکارید مجزا به نام آمیلوز و آمیلوپکتین هستند که نسبت آنها بسته به منبع نشاسته متفاوت است.
- آمیلوز یک پلیمر غیر شاخهای گلوکز 1→4 است که میتواند دارای طول چند هزار واحد گلیکوزیدی باشد. زنجیره پلی ساکارید یک مارپیچ چپگرد با شش باقیمانده گلیکوزیدی در هر دور است. اعتقاد بر این است که رنگ آبی معروف حاصل از واکنش نشاسته با ید، ناشی از هماهنگی یونهای ید درون مارپیچ است. (در واقع، چنین واکنش رنگی زمانی رخ میدهد که مارپیچ دارای حداقل شش چرخش مارپیچی یا 36 واحد گلیکوزیدی باشد
- آمیلوپکتین شباهت زیادی به گلیکوژن دارد و یک پلی ساکارید (قند مرکب) با تعداد زیادی واحد گلوکزیل (قند مرتبط با گلوکز) با پیوند ۱ به ۴ و تعداد کمی با پیوند ۱ به ۶ است.
گلیکوژن
- گلیكوژن، كه اغلب به عنوان نشاسته حیوانی شناخته می شود، یک هوموپلی ساکارید (قند مرکب با واحدهای یکسان) شاخهدار و ذخیرهکننده مواد مغذی است که در تمام سلولهای پستانداران، برخی از پروتوزوآها و جلبکها یافت میشود.
- این ماده به ویژه در سلولهای کبد و ماهیچه انسان و سایر حیوانات شایع است. گلیکوژن نسبت به نشاسته قابلیت حل شدن بیشتری دارد و به صورت گرانولهای ریز در سیتوپلاسم (مایع درون سلولی) قرار دارد.
- بزرگترین مولکولهای گلیکوژن حاوی دهها هزار واحد گلوکز یا گلوسیل (برای مثال ۳۰،۰۰۰ واحد) هستند.
- هر مولکول گلیکوژن از زنجیرههای بلند و بسیار شاخهدار مولکولهای گلوکز تشکیل شده است.
- پیوندهای گلیکوزیدی (اتصالات بین قندها) بین کربنهای ۱ و ۴ گلوکز (یعنی اتصالات ۱ به ۴) تشکیل میشوند، به استثنای محلهای انشعاب که شامل اتصالات ۱ به ۶ هستند.
- یک مولکول گلیکوژن از سه نوع زنجیره متمایز تشکیل شده است: A، B و C. یک زنجیره C منفرد وجود دارد که حاوی چندین زنجیره B و A است و به یک گروه کاهنده آزاد (یعنی کربن ۱ گلوکز در انتهای زنجیره C که دارای یک گروه هیدروکسیل یا OH است) ختم میشود.
- زنجیرههای B مستقیماً به زنجیره C متصل شده و حداقل یک زنجیره A را حمل میکنند. علاوه بر این، زنجیره A میتواند به زنجیره C متصل شود.
سلولز
- سلولز فراوانترین و گستردهترین ماده زیستی روی کره زمین است. این ماده جزئی ضروری از دیواره سلولی گیاهان و همچنین دیواره سلولی جلبکها و قارچها است.
- سلولز یک پلیساکارید (قند مرکب) ساختاری غیرشاخهدار (صاف) از گلوکز (قند ساده) است که در آن مونوساکاریدهای (قندهای ساده) مجاور از طریق پیوندهای گلیکوزیدی β-۱ به ۴ به هم متصل میشوند.
- طول زنجیرههای سلولز از چند صد تا چند هزار واحد گلوسیل (قند مرتبط با گلوکز) متغیر است (برای مثال، در جلبک والونیا، یک مولکول سلولز ممکن است حاوی بیش از ۲۰،۰۰۰ واحد گلوسیل باشد).
- در مولکولهای سلولز، حلقههای پیرانوز (نوعی ساختار قندی) متوالی نسبت به یکدیگر ۱۸۰ درجه چرخیده میشوند و ظاهری زیگزاگی به زنجیره قند میدهند.
- به همین دلیل، گروههای OH مولکولهای قند از زنجیره در تمام جهات بیرون زده و به آنها اجازه میدهند تا پیوندهای هیدروژنی با گروههای OH زنجیرههای سلولز مجاور تشکیل دهند و در نتیجه یک شبکه سه بعدی ایجاد کنند.
- بنابراین، در دیواره سلولی گیاهان، ۲۰۰۰ مولکول سلولز به صورت موازی، ریز رشتههای متصل به هم (با قطر ۲۵ نانومتر) سازماندهی میشوند که محور بلند آنها همان محور زنجیره گلوکز منفرد است.
کیتین
- کیتین پلیمری است که در دیواره سلولی هیفهای قارچی و اسکلت خارجی بندپایان یافت میشود.
- ساختار شیمیایی کیتین شبیه به سلولز است، با این استثنا که گروه هیدروکسیل هر اتم کربن در موقعیت ۲ با یک گروه استیل آمید جایگزین میشود.
- بنابراین کیتین یک پلیمر غیرشاخهدار از N-استیل گلوکزآمین (قند با گروه آمینه) است که حاوی هزاران واحد آمین قند است که توسط پیوندهای گلیکوزیدی β-۱ به ۴ به هم متصل شدهاند.
- علاوه بر نشاسته و سلولز، سلولهای گیاهی حاوی سایر پلیساکاریدها مانند زایلان، اسیدهای آلژینیک (جلبکها)، اسیدهای پکتیک، اینولین، آگارآگار و همی سلولز هستند.
- برخی از این پلیساکاریدها برای سلول تکیهگاه مکانیکی فراهم میکنند، در حالی که برخی دیگر به عنوان ذخایر غذایی عمل میکنند.
۲. هتروپلی ساکاریدها (قندهای مرکب ناهمگن)
هتروپلی ساکاریدها پلی ساکاریدهایی (قندهای مرکب) هستند که در مولکولهای خود انواع مختلفی از مونوساکاریدها (قندهای ساده) و اسیدهای آمین دار، سولفوریک یا فسفریک دارند. مهمترین هتروپلی ساکاریدها عبارتند از:
- اسید هیالورونیک، سولفات کراتین و سولفات کندروتین
اسید هیالورونیک، سولفات کراتین و سولفات کندروتین:
اسید هیالورونیک، سولفات کراتین و سولفات کندروتین، هتروپلی ساکاریدهای اسیدی هستند که در بافت غضروف یافت می شوند.
- اسید هیالورونیک یک هتروپلی ساکارید غیرشاخهدار است که از دی ساکاریدهای تکرارشونده N-استیل گلوکزآمین یا D-گلوکزآمین و اسید گلوکورونیک تشکیل شده است.
- اسید هیالورونیک همچنین در سایر بافتهای همبند، مایع سینوویال مفاصل، زجاجیه چشم و کپسولهای باکتریایی وجود دارد.
- سولفات کراتین، مانند اسید هیالورونیک، یک دی ساکارید تکرارشونده است که یک زنجیره مستقیم ایجاد میکند. هر واحد دی ساکارید این پلی ساکارید از D-گالاکتوز و N-استیل گلوکزآمین سولفاته تشکیل شده است. این ماده جزئی از غضروف و قرنیه است.
- سولفات کندروتین یک دی ساکارید است که از باقی ماندههای متناوب اسید گلوکورونیک و N-استیل گالاکتوزآمین سولفاته تشکیل شده است. در غضروف، استخوان، پوست، ) میله غضروفی که در پشت جانوران است( notochord، آئورت و بند ناف وجود دارد.
هپارین
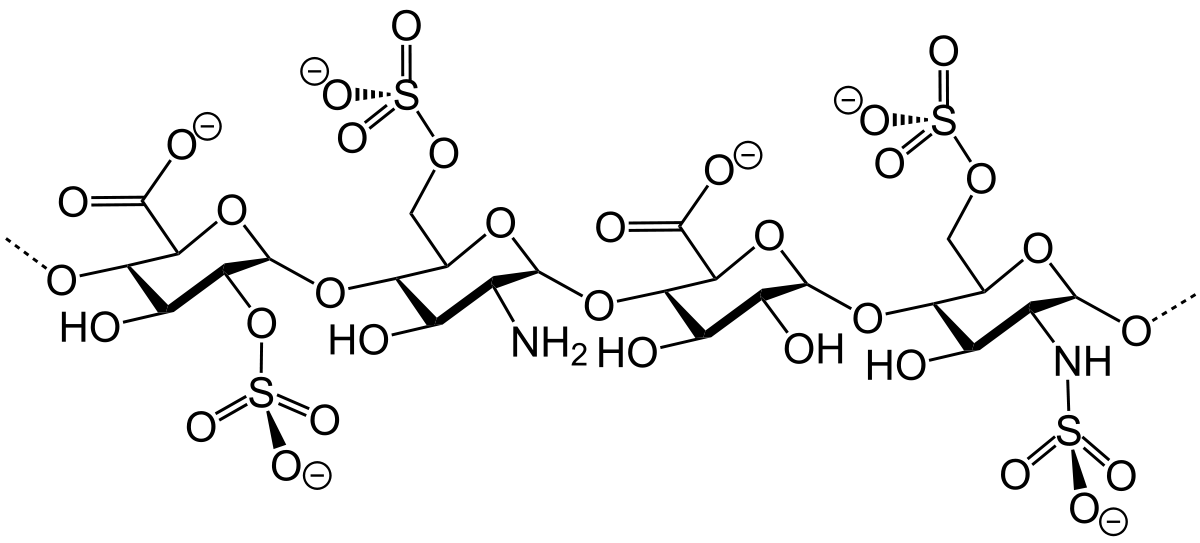
هپارین یک ضدانعقاد است که در خون، پوست، کبد، ریه، تیموس و طحال یافت می شود. مولکول آن از واحدهای دی ساکارید تکراری D-گلوکورونیک اسید و D-گلوکزآمین تشکیل شده است.
پروتئوگلیکان ها، گلیکوپروتئین ها و گلیکولیدها
علاوه بر این، پلی ساکاریدها به طور کووالانسی با پروتئین ها و لیپیدها ترکیب می شوند تا سه نوع مولکول زیر را تولید کنند:
- پروتئوگلیکان ها: مولکولهای پروتئوگلیکان عمدتا از پلی ساکاریدها و مقدار کمی پروتئین تشکیل شدهاند. همچنین به عنوان موکوپروتئینها شناخته میشوند. پروتئوگلیکانها آمورف هستند و ژلهایی تولید میکنند که قادر به حفظ مقادیر زیادی آب هستند. غضروف و استخوان شامل پروتئوگلیکان غضروفی خارج سلولی هستند.
- یک مولکول مرکزی طولانی از اسید هیالورونیک توسط رشتههای پروتئین، به نام پروتئین هسته، احاطه شده است. هر رشته پروتئین هسته حاوی سه بخش حاوی کربوهیدرات است. بخش اول عمدتا از الیگوساکاریدها، بخش دوم از زنجیرههای سولفات کراتین و بخش سوم از زنجیرههای سولفات کندروتین تشکیل شده است. این پیکربندی به غضروف مقاومت و استحکام کششی می دهد.
- گلیکوپروتئین ها (یا گلیکوزآمینوگلیکان ها یا موکوپلی ساکاریدها)
گلیکوپروتئینها مولکولهایی هستند که از ترکیب پروتئین و قند (پلی ساکارید) ساخته شدهاند. در این ترکیبات، زنجیره قندی به مراتب کوتاهتر و اغلب شاخهدار است. بسیاری از آنزیمها، هورمونها، گروههای خونی، اجزای بزاق دهان، مخاط معده، آلبومینهای خون، آنتیبادیها و ایمونوگلوبولینها گلیکوپروتئین هستند که نقشهای متنوعی را در سلولها و اندامها ایفا میکنند.
- گلیکول لیییدها
این مولکولها از اتصال کووالانسی (پیوند قوی) قند و چربی (لیپید) تشکیل شدهاند. بخش قندی میتواند شامل یک قند ساده (مونوساکارید) یا یک زنجیره خطی یا شاخهدار باشد. بخش عمدهای از غشای سلولی از گلیکولיפیدها، مانند سربروزیدها و گانگلیوزیدها، ساخته شده است.
۳. چربی ها (لیپیدها)
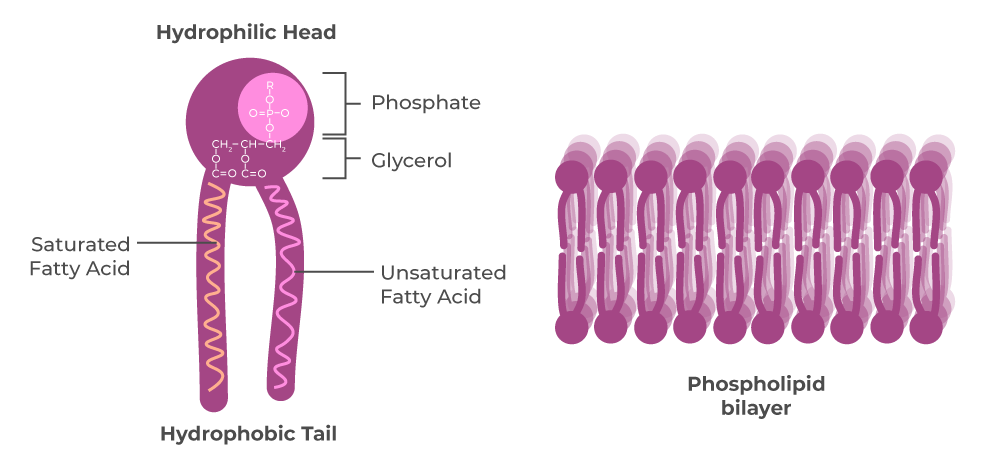
- چربیها (لیپیدها) که از کلمه یونانی lipos به معنای چربی گرفته شدهاند، در آب نامحلول هستند اما در حلالهای آلی غیرقطبی مانند استون، بنزن، کلروفرم و اتر محلول هستند.
- این ویژگی کلی لیپیدها به دلیل وجود زنجیرههای بلند هیدروکربنهای آلیفاتیک یا حلقههای بنزن در مولکولهای آنها است. به عبارت دیگر، لیپیدها آبگریز (دفع کننده آب) و غیرقطبی هستند.
- نمونههایی از لیپیدهای رایج شامل روغن نباتی، کره، روغن حیوانی، موم، لاستیک طبیعی و کلسترول است.
- شبیه به کربوهیدراتها، لیپیدها نیز دو عملکرد اصلی در سلولها و بافتها دارند:
- ساختمان غشای سلولی: لیپیدها به همراه پروتئینها اجزای اصلی غشای سلولی هستند که از سلول در برابر محیط خارجی محافظت میکند و کنترل میکند که چه مواد میتوانند وارد یا خارج شوند.
- ذخیره انرژی: چربیها میتوانند به عنوان منابع ذخیره انرژی درون سلولها نگهداری شوند. مشابه نشاسته و گلیکوژن، چربی به دلیل فشردگی و نامحلول بودن در آب، ساختار مفیدی برای ذخیره مولکولهای تولید کننده انرژی (اسیدهای چرب) برای استفاده در مواقع نیاز فراهم میکند.
لیپیدها از کربن، هیدروژن و گاهی اوقات اکسیژن تشکیل شدهاند. در مقایسه با تعداد اتمهای کربن، تعداد اتمهای اکسیژن در مولکولهای چربی همیشه ناچیز است. گاهی اوقات فسفر، نیتروژن و گوگرد نیز به مقدار کم وجود دارند.
چربیها و روغنهای طبیعی از گلیسرول (گلیسیرین یا پروپان ۱، ۲، ۳-تریول) و مولکولهای اسید چرب تشکیل شدهاند.
این مواد استرهایی هستند که از ترکیب اسیدهای آلی با الکلها به وجود میآیند. تنها یک نوع گلیسرول وجود دارد که ساختار مولکولی آن در همه لیپیدها یکسان است و فرمول آن C3H8O3 و ساختار شیمیایی زیر را دارد:
اسیدهای چرب: مولکول اسید چرب آمفیدار است (دارای دو سر مجزا) و از دو بخش یا انتهای جداگانه تشکیل شده است:
- یک زنجیره هیدروکربنی بلند که آبگریز (نامحلول در آب) و از نظر شیمیایی غیرفعال است.
- یک گروه کربوکسیلیک اسید که در محلول یونیزه میشود (COO-)، به شدت آبدوست (محلول در آب) است و به سرعت استر و آمید تولید میکند.
نمکهای اسید چرب در مایعات خنثی، قطرات کروی کوچک یا میسلهایی را با گروههای کربوکسیل جدا شده در سطح و زنجیرههای کربنی آبگریز که به سمت مرکز امتداد مییابند، تشکیل میدهند. در سلولها، اسیدهای چرب به سایر اجزا استری شده و لیپیدهای قابل صابونی شدن تولید میکنند. اسیدهای چرب به صورت آزاد نادر هستند و برای ایجاد لیپیدهای قابل صابونی شدن، استری میشوند.
یک مولکول اسید چرب میتواند اشباع شده یا غیر اشباع باشد.
- با زنجیرههای هیدروکربنی بلند که به یک گروه کربوکسیل انتهایی ختم میشوند، مشخص میگردند.
- این اسیدها با فرمول کلی CH3 – (CH2)n – COOH مطابقت دارند.
- در اکثر اسیدهای چرب طبیعی، n یک عدد زوج بین 2 و 22 است.
- در اسیدهای چرب اشباع که بیشتر در بافتهای حیوانی یافت میشوند، n معمولاً ۱۲ (اسید میریستیک)، ۱۴ (اسید پالمیتیک) یا ۱۶ (اسید استئاریک) است.
اسیدهای چرب غیراشباع
- در اسیدهای چرب غیراشباع، حداقل دو تا، اما به طور معمول بیش از شش عدد از اتمهای کربن زنجیره هیدروکربنی توسط پیوندهای دوگانه (- C = C -) به هم متصل میشوند (به عنوان مثال، اسیدهای اولئیک، لینولئیک، لینولنیک، آراشیدونیک و کلپانوئیدونیک).
- وجود پیوندهای دوگانه در این اسیدها باعث افزایش انعطافپذیری زنجیره هیدروکربنی و در نتیجه سیالیت غشای زیستی میشود.
- اسیدهای چرب غیراشباع در لیپیدهای گیاهان عالی و جانورانی که با سرما سازگار شدهاند، غالب هستند.
- مقادیر بیشتری از اسیدهای چرب اشباع را میتوان در لیپیدهای حیواناتی که در مناطق گرمتر زندگی میکنند یافت.
اسیدهای چرب ضروری
- موجودات خاص، به ویژه پستانداران، قادر به سنتز برخی از اسیدهای چرب نیستند و بنابراین به آنها در رژیم غذایی خود نیاز دارند.
- اسیدهای چرب ضروری شامل اسید لینولئیک، اسید لینولنیک و اسید آراشیدونیک هستند.
- این اسیدهای چرب ضروری باید توسط حیوان از مواد گیاهی به دست آیند.
انواع لیپیدها
لیپیدها به سه دسته اصلی طبقه بندی می شوند:
- لیپیدهای ساده
- لیپیدهای مرکب
- لیپیدهای مشتق شده
- لیپیدهای ساده:
لیپیدهای ساده استرهای الکل با اسیدهای چرب هستند. این دسته خود به دو زیرگروه تقسیم می شوند:
- چربی های خنثی (گلیسیریدها یا تری گلیسریدها): گلیسیریدها و تری گلیسریدها استرهای سه گانه اسیدهای چرب و گلیسرول هستند. بخش عمده ای از چربی های ذخیره شده، چربی های خنثی هستند که در سیتوپلاسم سلول تجمع می یابند.
- موم ها: موم ها نسبت به چربی های خنثی نقطه ذوب بالاتری دارند و استرهای اسیدهای چرب با وزن مولکولی بالا با الکل هایی به غیر از گلیسرول هستند. کلسترول مهمترین الکل موجود در مولکول های موم هایی مانند موم زنبور عسل است.
- لیپیدهای مرکب:
در مولکول های لیپیدهای مرکب علاوه بر اسیدهای چرب و الکل، ترکیبات دیگری مانند فسفر، نیتروژن آمینی، کربوهیدرات ها و غیره وجود دارد. برخی از لیپیدهای مرکب اجزای ساختاری ضروری سلول به ویژه غشای سلولی هستند. لیپیدهای مرکب سلولی از انواع زیر هستند:
(الف) فسفولیپیدها (یا گلیسروفسفو-فتیت ها):
این لیپیدها جزء اصلی غشای سلولی هستند. در مولکول فسفولیپید، دو گروه هیدروکسیل (OH) در گلیسرول به اسیدهای چرب متصل شده اند، در حالی که سومین گروه هیدروکسیل (OH) به اسید فسفریک متصل است. فسفات همچنین به یک مولکول دوست آب مانند اتانول آمین، کولین، اینوزیتول یا سرین متصل می شود. بنابراین، هر مولکول فسفولیپید شامل یک دم آبگریز (نامحلول در آب) ساخته شده از دو زنجیره اسید چرب و یک سر قطبی آب دوست (محلول در آب) حاوی فسفات است.
در نتیجه، مولکول های فسفولیپید مانند مواد شوینده عمل می کنند. یعنی زمانی که مقدار کمی فسفولیپید روی سطح آب پخش شود، یک لایه نازک از مولکول های فسفولیپید تشکیل می شود. در این لایه نازک، بخش های دمی مولکول های فسفولیپید به صورت فشرده در برابر هوا قرار می گیرند، در حالی که سرهای آن ها با آب در تماس هستند.
دو لایه از این فیلمهای نازک میتوانند انتها به انتها ادغام شوند و یک ساندویچ فسفولیپیدی یا دولایه لیپیدی خود-درزگیر را تشکیل دهند که اساس ساختاری غشای سلولی است. غشای سلولی حاوی چهار نوع فسفولیپید است که در زیر فهرست شده اند:
- فسفاتیدیل کولین
- فسفاتیدیل اتانول آمین
- فسفاتیدیل سرین
- فسفاتیدیل اینوزیتول
سایر فسفولیپیدهای مهم ماتریکس شامل فسفوinositides، پلاسمالوژن ها و ایزوزیتیدها هستند. فسفوinositides به طور عمده در سلول های کبد، مغز، ماهیچه و سویا یافت می شوند. پلاسمالوژن ها یک خانواده تخصصی از فسفولیپیدها هستند که به ویژه در غشای سلول های عصبی و عضلانی و همچنین سلول های سرطانی فراوان هستند.
لیپوزوم ها:
هنگامی که سوسپانسیون های فسفولیپید آبی به سرعت با امواج فراصوت (یعنی سونوگرافی) هم زده شوند، لیپید در آب پراکنده می شود و لیپوزوم یا وزیکول های چربی را تشکیل می دهد. لیپوزوم ها اجسام کروی کوچکی (به قطر 25 نانومتر تا 1 میکرومتر) با دولایه ای از مولکول های فسفولیپید هستند که حجم کمی از محیط آبی اطراف را در بر می گیرند.
آنها بسیاری از ویژگی های نفوذپذیری غشاهای طبیعی را نشان می دهند، یعنی مولکول های ریز یا یون های محلول در آب می توانند توسط لیپوزوم ها محصور شوند و همچنین می توانند از دولایه لیپیدی آن عبور کنند.
به تازگی نشان داده شده است که لیپوزوم ها پتانسیل درمانی قابل توجهی را ارائه می دهند، زیرا می توانند به عنوان حامل برای انتقال داروهای خاص، پروتئین ها، هورمون ها، اسیدهای نوکلئیک، یون ها یا هر نوع مولکول دیگری به انواع خاصی از سلول های حیوانی مورد استفاده قرار گیرند.
دو مسیر به محتویات لیپوزوم اجازه دسترسی به سلول های هدف را می دهد:
- لیپوزوم ها می توانند به سطح سلول های هدف متصل شوند و با غشای پلاسمایی ادغام شوند و محتویات خود را به سیتوزول یا ماتریکس سیتوپلاسمی آزاد کنند.
- لیپوزوم ها می توانند به طور کامل توسط سلول بلعیده شده و در داخل سلول از بین بروند.
(ii) اسفنگول (Sphingolipids):
اسفنگولها به طور عمده در سلولهای مغزی یافت میشوند. مولکولهای آنها به جای گلیسرول، حاوی الکل آمینه (اسفینگول یا اسفنگوزین) هستند.
به عنوان مثال، غلافهای میلین فیبرهای عصبی شامل اسفنگومیلین هستند، لیپیدی که مولکولهای آن از اسفنگوزین و فسفولیپیدها تشکیل شدهاند.
(iii) گلیکولـیپیدها (Glycolipids):
مولکولهای گلیکولـیپیدها شامل هر دو کربوهیدرات و لیپید هستند. ماتریکس سلول حیوانی حاوی دو نوع گلیکولـیپید است:
- سربرُوزیدها (Cerebrosides): مولکولهای سربرُوزیدها حاوی اسفنگوزین، اسیدهای چرب و گالاکتوز یا گلوکز هستند. سربرُوزیدها اجزای ضروری ماده سفید سلولهای مغز و غلاف میلین اعصاب هستند. کراتین، سربرون، نروون و اوکسینروونها، سربرُوزیدهای مهمی هستند.
- گنگلیوزیدها (Gangliosides): گنگلیوزیدها از اسفنگوزین، اسیدهای چرب و یک یا چند مولکول گلوکز، لاکتوز، گالاکتوزآمین و اسید نورآمینیک تشکیل شدهاند. گنگلیوزیدها در ماده خاکستری مغز، غشای گلبولهای قرمز و سلولهای طحال یافت میشوند. گنگلیوزیدها دارای خواص آنتی ژنی هستند. گنگلیوزید GM2 ممکن است به دلیل نقص ژنتیکی که از تولید آنزیم تجزیهکننده این گنگلیوزید توسط سلولها جلوگیری میکند، در لیزوزومهای سلولهای مغزی تجمع یابد. این اختلال که به بیماری تای-ساکس (Tay-Sachs) معروف است، باعث فلج، کوری و تأخیر در رشد انسان میشود.
۴. لیپیدهای مشتق شده (یا لیپیدهای غیر قابل صابونی شدن)
برخی از انواع لیپیدها حاوی اسید چرب به عنوان جزء سازنده خود نیستند و این سه نوع عبارتند از:
الف) ترپن ها (Terpenes):
- ترپن ها شامل برخی ویتامین های محلول در چربی (مانند ویتامین های A، E و K)، کاروتنوئیدها (مانند رنگدانه های فتوسنتزی گیاهان) و برخی کوآنزیم ها (مانند کوآنزیم Q یا یوبیکینون) هستند.
- تمام ترپن ها از تعداد متفاوتی واحد ایزوپرن، یک واحد سازنده پنج کربنه، ساخته می شوند. واحدهای ایزوپرن به صورت سر به دم به هم متصل می شوند.
- دو مولکول ایزوپرن با هم ترکیب شده و یک مونوترپن را تشکیل می دهند، چهار واحد یک دیترپن، شش واحد یک تریترپن و به همین ترتیب. مونوترپن ها مسئول رایحه و طعم های متمایز گیاهان هستند (به عنوان مثال، ژرانیول از گل شمعدانی، منتول از نعناع و لیمونن از لیمو).
- دولیکول فسفات برای انتقال قندهای فعال شده در تولید گلیکوپروتئین ها و برخی پلی ساکاریدها در غشاء سلولی استفاده می شود. دولیکول فسفات یک پلی ایزوپرنوئید (یعنی یک پلیمر زنجیره بلند از ایزوپرن) است.
- کاروتنوئیدها ترکیبات لیپیدی هستند که رنگدانه های سلول های حیوانی و گیاهی را تشکیل می دهند. حدود 70 کاروتنوئید در هر دو نوع سلول وجود دارد.
- از جمله کاروتنوئیدهای مهم سلولی می توان به α، β و γ کاروتن، رتینن، زانتوفیل ها، لاکتوفلاوین در شیر، ریبوفلاوین (ویتامین B2)، زانتوسيانين ها، کوآنزیم Q، آنتوسیانین ها، فلاون ها، فلاونول ها و فلاونون ها اشاره کرد. از نظر شیمیایی، همه کاروتنوئیدها ایزوپرنوئیدهایی با زنجیره های بلند و سری های متناوب از پیوندهای دوگانه هستند.
- آنها توسط بافت های گیاهی تولید می شوند و در لامل های کلروپلاست قرار می گیرند تا به جذب نور در طی فتوسنتز کمک کنند. در سلول های حیوانی، کاروتنوئیدها به عنوان پیش سازهای ویتامین A عمل می کنند.
ب) کلروفیل ها (Chlorophylls):
- کلروفیل ها رنگدانه های فتوسنتزی سبز حیاتی کلروپلاست ها هستند. یک مولکول کلروفیل دارای یک سر و یک دم است.
- سر از یک حلقه پورفیرین یا هسته تتراپیرول تشکیل شده است که از آن یک دم آبگریز به نام فیتول که یک خوشه 20 کربنه است، امتداد می یابد.
- فیتول (C20H39) یک الکل با زنجیره مستقیم و یک پیوند دوگانه است. این را می توان شکلی از کاروتن (ویتامین A) با هیدروژن اضافی در نظر گرفت.
- پورفیرین ها (یونانی: porphyra = ارغوانی) مولکول های پیچیده کربن-نیتروژن هستند که به طور معمول یک فلز را احاطه می کنند، یعنی از چهار حلقه پیرول که توسط پل های متان به هم متصل شده اند و یک اتم فلز (منیزیم یا آهن) به حلقه های پیرول متصل است، تشکیل شده اند.
- پورفیرین در مولکول کلروفیل یک یون منیزیم را احاطه می کند، اما در هم مولکول هموگلوبین یک یون آهن را احاطه می کند. بسیاری از رنگدانه های اضافی سلول های حیوانی مانند میوگلوبین و سیتوکروم ها نیز دارای حلقه های پورفیرین در مولکول های خود هستند.
ج) استروئیدها (Steroids):
- استروئیدها از یک سیستم حلقههای سیکلوهگزان و سیکلوپنتان ادغام شده تشکیل شدهاند. همه آنها مشتقاتی از پرهیدرو-سیکلوپنتانو-فنترن هستند که از سه حلقه سیکلوهگزان ادغام شده و یک حلقه سیکلوپنتان انتهایی تشکیل شده است.
- استروئیدها خواص فیزیولوژیکی بسیار متفاوتی دارند. برخی از استروئیدها، به عنوان مثال، هورمون هستند (مانند هورمون های جنسی مانند استروژن، پروژسترون، تستوسترون و کورتیکوسترون) و بر بیان ژن برای تغییر عملکرد سلولی تأثیر می گذارند. برخی از استروئیدها ویتامین هستند (مانند ویتامین D2) و فعالیت آنزیم های خاصی را در سلول ها تنظیم می کنند. برخی از استروئیدها مانند اسید چولیک، امولسیفایرهای چربی صفرا هستند. استرول ها الکل های استروئیدی هستند. کلسترول موجود در حیوانات و ارگوسترول و استیگماسترول موجود در گیاهان نمونه های معمولی از استرول ها هستند.
- سلول های حیوانی زیادی کلسترول را در غشای پلاسمایی خود و همچنین در خون، صفرا، سنگ صفرا، مغز، نخاع و غدد فوق کلیوی دارند. کلسترول پیش سازه اکثر هورمون های جنسی استروئیدی و کورتیزون ها است.
- 7-دهیدروکلسترول در پوست وجود دارد که در حضور نور خورشید مسئول تولید ویتامین D است. علاوه بر این، ارگوسترول پیش ساز ویتامین D است.
د) پروستاگلاندین ها (Prostaglandins):
- پروستاگلاندین ها مشتقات هیدروکسی اسیدهای چرب چند غیر اشباع با 20 کربن هستند. مایع منی، بیضه، کلیه، جفت، رحم، معده، ریه، مغز و قلب انسان حاوی آنها هستند.
- حداقل شانزده پروستاگلاندین متمایز وجود دارد که در نه دسته (PGA، PGB، PGC … PGI) طبقه بندی می شوند. هدف اصلی آنها اتصال هورمون ها به غشای سلول های هدف است. پروستاگلاندین ها به طور مداوم در غشاها از پیش سازه هایی که توسط فسفولیپازها از فسفولیپیدهای غشایی شکسته می شوند، تشکیل می شوند. پروستاگلاندین ها میانجی های شیمیایی هستند که به صورت موضعی تولید می شوند.
- آغاز انقباض عضلات صاف (بنابراین به زایمان کمک می کند)، تجمع پلاکت ها و التهاب (یعنی آرتریت) از دیگر فعالیت های ضروری آنها هستند.
۵. پروتئین ها (Proteins)

- پروتئین ها از نظر شیمیایی و فیزیکی، متنوع ترین مولکول های درشت (ماکرومولکول) در سلول هستند.
- بیش از پنجاه درصد از وزن خشک سلول را این اجزای ضروری تشکیل می دهند.
- کلمه پروتئین توسط شیمی دان هلندی G.J. مولدر (1802-1880) ابداع شد و از کلمه یونانی proteios به معنای “در بالاترین رتبه” گرفته شده است. پروتئین ها جزء اصلی ساختاری پروتوپلاسم هستند و نقش های حیاتی دیگری را در سیستم های بیولوژیکی ایفا می کنند.
- آنزیم ها پروتئین های تخصصی کروی شکلی هستند که به عنوان کاتالیزور در تقریباً تمام فرآیندهای بیوشیمیایی درون سلول ها عمل می کنند.
- پروتئین های دیگر شامل آنتی بادی ها (ایمونوگلوبولین ها)، پروتئین های حمل و نقل، پروتئین های ذخیره سازی، پروتئین های انقباضی و چندین هورمون هستند.
- در هر موجود زنده هزاران پروتئین متمایز وجود دارد که هر کدام دارای عملکرد ساختاری یا عملکردی خاصی هستند.
- یک سلول انسانی می تواند بیش از 10000 مولکول پروتئینی متمایز داشته باشد. پروتئین ها به لحاظ شیمیایی پلیمری از اسیدهای آمینه هستند.
۶. آنزیم ها (Enzymes)
- آنزیم ها که مواد شیمیایی بسیار مهمی هستند، در سیتوزول و چندین اندامک سلولی قرار دارند. آنزیم ها پروتئین های تخصصی هستند که قادر به عمل به عنوان کاتالیزور در واکنش های شیمیایی هستند.
- آنزیم ها مانند سایر کاتالیزورها در دنیای شیمی، کاتالیزورهای دنیای بیولوژیک هستند. آنها سرعت یک واکنش شیمیایی را تحت تاثیر قرار می دهند در حالی که در پایان واکنش عمدتا دست نخورده باقی می مانند.
- ماده ای که آنزیم روی آن کار می کند، سوبسترا (substrate) نامیده می شود. آنزیم ها نقش اساسی در بسیاری از فرآیندهای متابولیک و بیوسنتزی سلولی ایفا می کنند، از جمله سنتز (آنابولیسم) مولکول های DNA، RNA و پروتئین و تجزیه (کاتابولیسم) کربوهیدرات ها، لیپیدها و سایر ترکیبات شیمیایی.
طبقه بندی آنزیم های ماتریکس و اندامک های سلولی به شرح زیر است:
اکسیدورِدوکتازها (Oxireductases):
اکسیدورِدوکتازها آنزیم هایی هستند که واکنش های اکسیداسیون و احیا در سلول را کاتالیز می کنند. این آنزیم ها مانند هیدروژنازها یا ردوکتازها، اکسیدازها، اکسیژنازها و پراکسیدازها، الکترون ها و یون های هیدروژن را از سوبستراهای مربوطه خود انتقال می دهند.
ترانسفرازها (Transferases):
ترانسفرازها آنزیم هایی هستند که گروه های یک کربنه، آلدهیدی یا کتون دار، آسیل، گلیکوزیل، آلکیل، نیتروژنی، فسفر دار و گوگرد دار را از یک مولکول به مولکول دیگر منتقل می کنند.
هیدرولازها (Hydrolases):
این آنزیم ها با تزریق آب در سراسر پیوند شکسته شده، یک مولکول پیچیده را به دو مولکول هیدرولیز می کنند. پیوندهای زیر تحت تأثیر این آنزیم ها قرار می گیرند: استر، گلیکوزیل، اتر، پپتید، سایر پیوندهای C–N، انیدرید اسید، C–C، هالید و پیوندهای P–N. پروتئازها، استرازها، فسفاتازها، نوکلئازها و فسفوریلازها برخی از مهمترین آنزیم های هیدرولاز هستند.
لیازها (Lysases):
آنزیم های لیاز گروه هایی را به ترکیبات شیمیایی حاوی پیوند دوگانه اضافه یا حذف می کنند. پیوندهای C–C، C–O، C–N، C–S و C–هالید تحت تأثیر لیازها قرار می گیرند.
ایزومرازها (Isomerases):
این آنزیم ها ایزومریزاسیون یا بازآرایی درون مولکولی سوبستراها را کاتالیز می کنند، به عنوان مثال اکسیدورِدوکتازهای درون مولکولی، ترانسفرازهای درون مولکولی، لیازهای درون مولکولی، سیس-ترانس ایزومرازها، راسم آزها و اپی مرازها.
لی گازها یا سنتتازها (Ligases or Synthetases):
این آنزیم ها با شکستن پیوند فسفات، اتصال مولکول ها را کاتالیز می کنند. این آنزیم ها پیوندهای C–O، C–S، C–N و C–C ایجاد می کنند.
طبقه بندی آنزیم ها بر اساس ماهیت شیمیایی سوبسترا:
طبقه بندی دیگری از آنزیم ها بر اساس ماهیت شیمیایی سوبسترا وجود دارد: کربوهیدراتازها در رتبه اول قرار دارند، به دنبال آنها پروتئازها (اندوپپتیدازها و اگزوپپتیدازها)، آمیلازها، استرازها، دهیدروژنازها، اکسیدازها، دکربوکسیلازها، هیدرازها، ترانسفرازها و ایزومرازها قرار دارند.
عوامل متعددی از جمله pH، دما و غلظت سوبسترا بر سرعت فعالیت آنزیم تأثیر می گذارند. فعالیت آنزیم وابسته به سوبسترا است.
برخی از آنزیم ها به صورت پیش سازیم (پروآنزیم) یا زیموژن وجود دارند که توسط کینازها برای انجام فعالیت کاتالیزوری فعال می شوند. به طور مشابه، تریپسینوژن از سلول های پانکراس توسط انتروکیناز در روده فعال می شود و پپسینوژن از سلول های اصلی معده توسط اسید هیدروکلریک تولید شده توسط سلول های جداری فعال می شود.
گروه های مصنوعی و کوآنزیم ها (Prosthetic Groups and Coenzymes)
- برخی از آنزیم ها مانند سیتوکروم ها پروتئین های متصل شده ای هستند که گروه های مصنوعی مانند متالوپورفیرین ها را در مولکول های خود دارند.
- برخی از آنزیم ها بدون حضور مولکول های کوآنزیم نمی توانند عمل کنند.
- آپوآنزیم (Apoenzyme) به آنزیم غیرفعال که به تنهایی قادر به عملکرد نیست اشاره دارد. هولآنزیم (Holoenzyme) به ترکیب آپوآنزیم و کوآنزیم اشاره دارد.
- به عنوان مثال، آنزیم هیدروژناز یک آپوآنزیم است که می تواند با NAD+ یا NADP به عنوان کوآنزیم کار کند.
برخی از کوآنزیم ها یا کوفاکتورها ضروری عبارتند از:
- نیکوتین آمید آدنین دینوکلئوتید (NAD) یا دی فسفوپیریدین نوکلئوتید (DPN)
- نیکوتین آمید آدنین دینوکلئوتید فسفات (NADP) یا تری فسفوپیریدین نوکلئوتید (TPN)
- فلاوین آدنین مونونوکلئوتید (FAM)
- فلاوین آدنین دینوکلئوتید (FAD)
- یوبیکوینون (کوآنزیم Q یا کوآنزیم کیو)
- اسید لیپوئیک (LIP یا S2)
- آدنوزین تری فسفات (ATP) و پیریدوکسیل فسفات (PALP)
- آدنوزیل متیونین
- اسید تتراهیدروفولیک (CoF)
- آدنوزیل متیونین
- بیوتین، کوآنزیم A (CoA)، تیامین пироفسفات (TPP) و اوریدین دی فسفات (UDP)
- ایزوآنزیم ها (Isoenzymes)
تحقیقات اخیر نشان داده است که چندین آنزیم عملکردهای مشابه و ساختار شیمیایی تقریباً یکسانی دارند.
به این آنزیم ها ایزوآنزیم گفته می شود. ایزوآنزیم ها با توارث ارتباط دارند.
در یک سلول حدود 100 ایزوآنزیم مانند پنج شکل یکسان از لاکتات دهیدروژناز (LDH) وجود دارد.
ویتامین ها (Vitamins)
- ویتامین ها مولکول های آلی با ساختار شیمیایی متنوع هستند. آنها برای رشد، عملکرد و تولید مثل طبیعی سلول به مقادیر کم نیاز هستند.
- ویتامین ها نقش مهمی در متابولیسم سلولی دارند و به عنوان آنزیم یا سایر کاتالیزورهای بیولوژیکی در بسیاری از فرآیندهای شیمیایی سلول عمل می کنند.
- هپکینز، اوزبورن، مندل و مک کالوم اهمیت آنها را برای حیوانات گزارش کرده اند (1912-1913).
- فونک (1912) وجود بازهای نیتروژنی را در آنها ثابت کرد و به آنها اصطلاح “ویتامین” داد که به آمین های حیاتی اشاره دارد.
- سلول حیوانی قادر به ساخت ویتامین ها از رژیم غذایی معمولی نیست، بنابراین باید همراه با غذا مصرف شوند. کمبود آنها در سلول ها منجر به اختلال عملکرد متابولیک و بیماری های متعدد می شود.
- به عنوان مثال، کمبود اسید آسکوربیک (ویتامین C) تولید مارپیچ پروکلاژن را مختل می کند. کلاژنازها که آنزیم های خارج سلولی خاصی هستند، به طور مداوم کلاژن های طبیعی را از بین می برند.
- در اسکوربوت، زنجیره های “پیش ساز” (pro-chain) که به درستی تولید نشده اند، قادر به تشکیل مارپیچ سه گانه نیستند و به سرعت از بین می روند.
- در نتیجه از دست دادن تدریجی کلاژن طبیعی در ماتریکس، رگ های خونی بیش از حد ضعیف می شوند و دندان ها از حفره های خود خارج می شوند.
- این نشان می دهد که تخریب و جایگزینی کلاژن در این بافت ها به خصوص سریع است. به عنوان مثال، در استخوان ها، “گردش” کلاژن بسیار کند است، به این معنی که مولکول های کلاژن قبل از حل شدن و جایگزینی حدود 10 سال باقی می مانند.
هورمون ها (Hormones)
- هورمون ها مولکول های شیمیایی پیچیده ای هستند که به مقدار کم در سیتوپلاسم وجود دارند و بر ایجاد RNA پیام رسان (mRNA)، آنزیم ها و سایر فرآیندهای فیزیولوژیکی درون سلولی تأثیر می گذارند.
- از مهم ترین هورمون ها می توان به هورمون رشد، استروژن، آندروژن، انسولین، تیروکسین، کورتیزون و غیره اشاره کرد.
- این هورمون ها توسط غدد بدون مجرا یا غدد درون ریز تولید می شوند و از طریق رگ های خونی به سلول های مختلف جانوران چند سلولی منتقل می شوند. در سلول ها، آنها فرآیندهای متابولیک متنوعی را کنترل می کنند.
- برای مثال، کشف شده است که هورمون اسدیسون(ecdysone) باعث ایجاد پف کردگی (حلقه های بالبیانی) روی کروموزوم های بزرگ حشرات می شود.
- هورمون ها ژن را در مکان خاص کروموزومی فعال یا غیرفعال می کنند. بنابراین، هورمون ها برای تنظیم اعمال متعددی که با یک عملکرد خاص مرتبط هستند، عمل می کنند. برای نمونه، هورموناس دیسون (ecdysone) ریختن پوست و دگردیسی حشرات را کنترل می کند.
- سلول های جزایر لانگرهانس در پانکراس، هورمون انسولین را تولید می کنند که آنزیم های تبدیل گلوکز به گلیکوژن را در سلول های کبدی پستانداران تنظیم می کند.
علاوه بر این، هورمون تیروکسین تیروئید، آنزیم فسفریلاز را برای تبدیل گلیکوژن به گلوکز فسفات تحریک می کند.
۷. اسیدهای نوکلئیک (Nucleic Acids)
- اسیدهای نوکلئیک مولکولهای شیمیایی پیچیده و درشتمولکولی هستند که اهمیت بیولوژیکی بسیار زیادی دارند. آنها فرآیندهای متابولیک ضروری سلول را تنظیم میکنند و اطلاعات ژنتیکی را از نسلی به نسل دیگر منتقل میکنند.
- در موجودات زنده، دو نوع اسید نوکلئیک وجود دارد: اسید ریبونوکلئیک (RNA) و اسید دئوکسیریبونوکلئیک (DNA). هر دو شکل اسیدهای نوکلئیک از پلیمرهای نوکلئوتید تشکیل شدهاند.
- اجزای یک نوکلئوتید، نوکلئوزید و اسید فسفریک هستند. حتی نوکلئوزید نیز از قندهای پنتوز (ریبوز یا دئوکسی ریبوز) و بازهای نیتروژنی (پورینها یا پیریمیدینها) تشکیل شده است.
- آدنین و گوانین پورین هستند، در حالی که سیتوزین، تیمین و اوراسیل پیریمیدین هستند. سیتوزول (محفظهی داخلی سلول) تنها از RNA تشکیل شده است، در حالی که DNA منحصراً در هسته متمرکز باقی میماند.
- DNA و RNA از نظر ساختار شیمیایی تقریباً یکسان هستند، با چند تفاوت جزئی.
خواص سیتوزول (Cytoplasmic Matrix)
ماتریکس یک ماده زنده است و دارای خواص فیزیکی و بیولوژیکی زیر است:
خواص فیزیکی
به دلیل ماهیت کلوئیدی ماتریکس، اکثر خواص فیزیکی آن به شرح زیر است:
۱. اثر تیندال (Tyndall’s effect)
وقتی در محیط تاریک، پرتو قوی نور به صورت عمود به سیستم کلوئیدی ماتریکس تابانده شود، نور توسط ذرات کلوئیدی ریز معلق در سیستم کلوئیدی منعکس می شود. مسیر نور به شکل مخروط دیده می شود. از آنجایی که تیندال (۱۸۲۰-۱۸۹۳) اولین کسی بود که این پدیده را در کلوئیدها توصیف کرد، این مخروط نور به عنوان مخروط تیندال شناخته می شود.
۲. حرکت براونی (Brownian movement)
ذرات کلوئیدی معلق در ماتریکس همیشه به صورت زیگزاگی حرکت می کنند. این حرکت مولکول ها بر اثر برخورد مولکول های آب ایجاد می شود که حرکت را به مولکول های کلوئیدی منتقل می کنند.
رابرت براون، گیاه شناس اسکاتلندی، برای اولین بار در سال ۱۸۲۷ این نوع حرکت را در محلول کلوئیدی کشف کرد. در نتیجه، این حرکات به عنوان حرکت براونی نامیده می شوند. حرکت براونی از ویژگی های تمام مایعات کلوئیدی است و به اندازه ذرات و دما وابسته است.
۳. سیتوپلاسم و حرکت آمیبی (Cyclosis and amoeboid movement)
به دلیل خاصیت تغییر فاز سیتوزول، جریان یا حرکت درون سلولی ماتریکس رخ می دهد. این ویژگی حرکتی ماتریکس درون سلولی به سیتوپلاسم (چرخش درون سلولی) معروف است. به طور معمول، سیتوپلاسم در فاز سول ماتریکس رخ می دهد و تحت تأثیر فشار هیدرواستاتیک، دما، PH، ویسکوزیته و غیره قرار می گیرد.
حرکات درون سلولی پینوزوم ها، فاگوزوم ها و اندامک های مختلف سیتوپلاسمی مانند لیزوزوم ها، میتوکندری ها، کروموزوم ها، سانتریول ها و غیره، تنها نتیجه سیتوپلاسم است. این پدیده در اکثر سلول های حیوانی و گیاهی مشاهده شده است. تحرک آمیب ها مستقیماً به سیتوپلاسم وابسته است.
حرکت آمیبی در پروتوزوآها، گلبول های سفید، سلول های اپیتلیال، مزانشیمی و سایر سلول ها رخ می دهد. در طی حرکت آمیبی، سلول به طور فعال شکل خود را تغییر می دهد و پزودوپودیا (پاهای کاذب) که برآمدگی های سیتوپلاسمی هستند را بیرون می فرستد. به دلیل سیتوپلاسم، ماتریکس این پزودوپودیاها را حرکت می دهد که سلول را به جلو هدایت می کند.
۴. کشش سطحی (Surface tension)
مولکول ها در داخل یک مایع همگن آزادانه حرکت می کنند و به طور مساوی توسط مولکول های همسایه از تمام جهات کشیده می شوند. در سطح مایع، جایی که با هوا یا مایع دیگری تماس پیدا می کند، مولکول ها به سمت پایین، طرفین یا داخل بیشتر از بالا کشیده می شوند؛ در نتیجه، تحت تنش های نابرابر قرار گرفته و برای ایجاد غشاء در کنار هم نگه داشته می شوند.
نیرویی که مولکول ها را در کنار هم نگه می دارد به عنوان کشش سطحی مایع شناخته می شود. سیتوزول به عنوان یک مایع، خاصیت کشش سطحی دارد. پروتئین ها و لیپیدهای ماتریکس کشش سطحی پایینی دارند، بنابراین در سطح قرار می گیرند و غشاء را تشکیل می دهند، در حالی که ترکیبات شیمیایی مانند NaCl کشش سطحی بالایی دارند و در داخل ماتریکس یافت می شوند.
۵. جذب سطحی (Adsorption)
جذب سطحی، افزایش غلظت یک ماده در سطح محلول است (L., ad=to, sorbex=to draw in).
جذب سطحی به ماتریکس در تشکیل مرزهای پروتئینی کمک می کند.
۶. سایر خواص مکانیکی یا فیزیکی ماتریکس
علاوه بر کشش سطحی و جذب سطحی، ماتریکس دارای انواع دیگر ویژگی های مکانیکی از جمله خاصیت ارتجاعی، انقباض پذیری، سفتی و ویسکوزیته است که مزایای فیزیولوژیکی متعددی را برای ماتریکس به ارمغان می آورد.
قطبیت تخم (Polarity of the egg)
به دلیل فاز پایدار سیستم کلوئیدی، قطبیت ماتریکس سلولی را نمی توان با سانتریفیوژ یا سایر روش های مکانیکی تغییر داد.
بافرها و PH (Buffers and pH)
ماتریکس دارای یک مقدار PH ثابت است و نمی تواند تغییرات قابل توجهی در تعادل PH خود را تحمل کند. با این حال، فرآیندهای متابولیک متعدد مقادیر کمی از اسیدهای اضافی یا بازها تولید می کنند. برای محافظت در برابر چنین نوسانات PH، ماتریکس حاوی بافرهایی مانند سیستم کربنات-بی کربنات است که سطح PH ثابتی را حفظ می کند.
خواص زیستی (Biological Properties)
ماتریکس یک ماده زنده است که دارای ویژگی های بیولوژیکی زیر است:
- تحریک پذیری (Irritability): تحریک پذیری ماتریکس یک ویژگی اساسی و ذاتی است. حساسیت به تحریک، توانایی انتقال برانگیختگی و توانایی پاسخ به محرک را دارد. انقباض سیتوزول توسط گرما، نور، ترکیبات شیمیایی و سایر عوامل تحریک می شود.
- هدایت پذیری (Conductivity): هدایت پذیری فرآیند هدایت یا انتقال برانگیختگی از منبع آن به منطقه واکنش آن است. ماتریکس سلول های عصبی خاصیت هدایت پذیری را نشان می دهد.
- حرکت (Movement): سیتوزول به دلیل سیتوپلاسم قادر به حرکت است. سیتوپلاسم به سن، میزان آب، عوامل ارثی و ترکیب سلولی بستگی دارد.
- متابولیسم (Metabolism): ماتریکس محل بسیاری از فرآیندهای شیمیایی درگیر در متابولیسم است. این فعالیت ها می توانند سازنده یا مخرب باشند. فعالیت های آنابولیک شامل ایجاد پروتئین ها، لیپیدها، کربوهیدرات ها و اسیدهای نوکلئیک است، در حالی که فرآیندهای کاتابولیک شامل اکسیداسیون غذا و سایر مواد است. اصطلاح ترکیبی برای فرآیندهای آنابولیک و کاتابولیک، فرآیند متابولیک است.
- رشد (Growth): به دلیل فعالیت های ترشحی یا آنابولیک سلول، حجم پروتوپلاسم تازه به طور مداوم افزایش می یابد. افزایش حجم ماتریکس منجر به بزرگ شدن سلول می شود که در نهایت با تقسیم سلولی به سلول های دختری تقسیم می شود.
- تولید مثل (Reproduction): سیتوپلاسم هم توانایی تولید مثل غیرجنسی و هم جنسی را دارد.
همچنین بخوانید:
- میکروفیلامنت چیست؟ تعریف، ساختار، عملکرد
- واکوئل چیست؟ ترکیب، ساختار، انواع و عملکرد
- اسکلت سلولی (Cytoskeleton): تعریف، ساختار، کارکردها و شکل آن
- ساختار های سلولی- دوبله ژنیران
- سانتریول چیست؟ تعریف، ساختار، کاربرد و تصاویر
- میکروتوبول (Microtubule): تعریف، ساختار، کارکردها و شکل آن
- سانتروزوم چیست؟ تعریف، ساختار، عملکرد، شکل
مترجم: محمد صادق محمودی لرد