


تکنیک ها
توالی یابی ایلومینا

توالی یابی ایلومینا : توالی یابی ایلومینا تکنیکی است که برای تعیین مجموعهای از جفت بازها در DNA استفاده میشود که به عنوان توالی یابی DNA نیز شناخته میشود. مفهوم شیمی برگشتپذیر و ناپذیر بودن واکنشها توسط Bruno Canard و Simon Sarfati در انستیتو پاستور پاریس ابداع شد. این روش توالی یابی مبتنی بر رنگهای برگشت پذیر است که شناسایی تک نوکلئوتیدها را در حین شسته شدن روی رشتههای DNA امکان پذیر میکند. همچنین میتواند برای توالی یابی کل ژنوم و توالی یابی بخشی از ژنوم، تجزیه و تحلیل رونویسی، متاژنومیکس، کشف RNA کوچک، پروفایل متیلاسیون، و تجزیه و تحلیل برهمکنش پروتئین-اسید نوکلئیک در سطح ژنوم استفاده شود.
چشم انداز:
DNA از طریق توالیهای مکمل به سلول مورد نظر می چسبد. رشته خم می شود و به اولیگوی دوم متصل می شود و یک پل را تشکیل میدهد. در ادامه پلیمراز، یک رشته معکوس را سنتز میکند. دو رشته آزاد شده و صاف میشوند. هر کدام یک پل جدیدی را تشکیل می دهند. در نتیجه مجموعهای از کلونهای رشتهای رو به جلو و معکوس DNA ایجاد میشود.
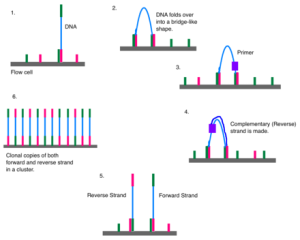
این تکنیک در سه مرحله اساسی خلاصه میشود: تکثیر، توالی یابی و تجزیه و تحلیل. این فرآیند با DNA خالص شده آغاز میشود. DNA قطعه قطعه شده و آداپتورهایی به این قطعهها متصل میشود که شامل بخشهایی میباشد که به عنوان نقاط مرجع در طول مراحل تکثیر، توالی یابی و تجزیه و تحلیل عمل میکنند. DNA اصلاح شده روی سلول هدف بارگذاری میشود که در آن تکثیر و توالی یابی انجام میشود. سلول اصلاح شده حاوی منافذ نانویی است که قطعات اضافی را از سلول خارج میکند و مانع از تجمع بیش از حد آنها در سلول میشوند. هر نانو منفذ حاوی الیگونوکلئوتیدهایی است که مانند یک نقطه اتصال برای متصل شدن به آداپتورها عمل میکند.
پس از اتصال قطعات، مرحلهای به نام سنتز رشتهای DNA آغاز میشود. این مرحله از هر قطعه DNA حدود هزار نسخه تهیه میکند و این کار توسط PCR انجام میشود. سپس آغازگرها و نوکلئوتیدهای اصلاح شده روی تراشهای شسته میشوند. این نوکلئوتیدها حالت برگشت پذیری دارند. DNA پلیمراز در هر بار فقط یک نوکلئوتید را همراه با مسدود کننده فلورسنت به قطعه DNA اضافه کند. پس از هر دور، دوربین از تراشه عکس میگیرد. کامپیوتر تعیین میکند که چه قطعهای با طول موج فلورسنت اضافه شده است و طول موج هر نقطه را روی تراشه ثبت میکند. پس از هر دور، مولکولهایی که به اشتباه متصل شدهاند، شسته میشوند. سپس یک مرحله شیمیایی برای حذف گروه مسدودکننده انتها فلورسنت 3′ وجود دارد. این روند تا زمانی ادامه مییابد که مولکول DNA به طور کامل توالی یابی شود. با استفاده از این فناوری، هزاران مکان در سراسر ژنوم به طور همزمان از طریق توالی یابی موازی گسترده، تعیین توالی میشوند.
روش:
کتابخانه ژنومیک
پس از خالص سازی DNA، یک کتابخانه DNA یا ژنومی، باید ایجاد شود. دو راه برای ایجاد یک کتابخانه ژنومی وجود دارد، Sonification و Tagmentation. با برچسب گذاری، transposases به طور تصادفی DNA را به قطعاتی بین 50 تا 500 جفت باز برش میدهند و همزمان آداپتورهایی را به قطعات ژنومی اضافه میکند. در یکی از کتابخانههای ژنتیکی عنوان شده است که میتوان از سونوگرافی برای قطعه قطعه کردن DNA ژنومی استفاده کرد. امواج فراصوت به همراه امواج صوتی اولتراسونیک میتواند DNA را به اندازههای مشابه قطعه قطعه کند. آداپتورهای راست و چپ باید توسط T7 DNA Polymerase و T4 DNA Ligase پس از فراصوت به رشته وصل شوند. رشتههایی که آداپتور به آنها متصل نمیشود، شسته میشوند.
آداپتورها:
آداپتورها شامل سه بخش مختلف هستند: توالی مکمل جایگاه هدف (الیگونوکلئوتیدها در سلول هدف)، توالی کد دار و محل اتصال برای متصل شدن به ناحیه آغازگر توالی. کدها معمولاً شش جفت باز هستند و در طول تجزیه و تحلیل توالی DNA برای شناسایی نمونهها استفاده میشوند. در طول تجزیه و تحلیل، رایانه همه خوانشها را با یک شاخص یکسان گروهبندی میکند. توالی یابی ایلومینا از رویکرد “توالی همراه با سنتز” استفاده میکند. این فرآیند در یک محفظه شیشهای با پوشش آکریل آمید انجام میشود. سلول مورد نظر حاوی الیگونوکلئوتیدهایی (توالیهای نوکلئوتیدی کوتاه) است، که این الیگونوکلئوتیدها کف سلول را میپوشاند و به عنوان تکیهگاه برای نگه داشتن رشتههای DNA در طول توالی یابی عمل میکنند. همانطور که DNA قطعه قطعه شده روی سلول مورد نظر شسته میشود، آداپتور مناسب به جایگاه مکمل متصل میشود.
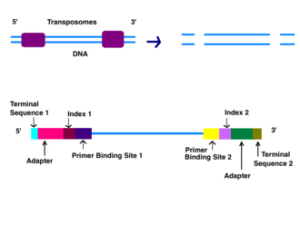
DNA دو رشتهای توسط ترانسپوزومها شکافته میشود. انتهای بریده شده ترمیم میشوند و آداپتورها، شاخص ها، محلهای اتصال پرایمر و محلهای پایانی به هر رشته DNA اضافه میشوند.
Bridge amplification:
پس از اتصال، تولید رشته DNA، آغاز میشود. هدف، ایجاد صدها رشته DNA یکسان است. برخی از رشتهها جهت سنتزشان به سمت جلو میباشد، به همین دلیل است که از آداپتورهای راست و چپ استفاده میشود. DNA پلیمراز در امتداد یک رشته DNA حرکت میکند و رشته مکمل آن را ایجاد میکند. رشته اصلی شسته میشود و تنها رشته معکوس باقی میماند. در بالای رشته معکوس یک دنباله آداپتور وجود دارد. رشته DNA خم میشود و به اولیگو که مکمل توالی آداپتور بالایی است، میچسبد. پلیمرازها به رشته معکوس متصل میشوند و رشته مکمل آن (که مشابه رشته اصلی است) ساخته میشود. اکنون DNA دو رشتهای دناتوره میشود تا هر رشته بتواند به طور جداگانه به یک توالی الیگونوکلئوتیدی متصل به سلول هدف متصل شود، یکی رشته معکوس، دیگری پیشرو. این فرآیند bridge amplification (ایجاد پل) نامیده میشود و برای هزاران رشته در سراسر سلول هدف به طور همزمان اتفاق میافتد.
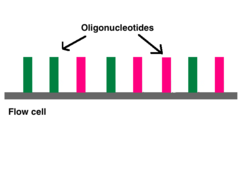
میلیونها الیگو در انتهای هر خط سلولی وجود دارد.
تکثیر کلونال:
بارها و بارها، رشتههای DNA خم میشوند و به تکیهگاه هدف متصل میشوند. DNA پلیمراز یک رشته جدید را برای ایجاد یک بخش دو رشتهای سنتز میکند و آن قطعه دناتوره میشود تا همه رشتههای DNA در یک ناحیه از دو منبع متفاوت باشند. تکثیر کلونال برای اهداف کنترل کیفیت اهمیت ویژهای دارد. اگر رشتهای دارای یک دنباله عجیب و غریب باشد، دانشمندان میتوانند رشته معکوس را بررسی کنند تا مطمئن شوند که مکمل همان رشته به چه شکل است. رشتههای رو به جلو و معکوس به عنوان کنترل برای محافظت در برابر نمونههای دیگر عمل میکنند. از آنجایی که توالی یابی Illumina از DNAپلیمراز استفاده میکند، خطاهای جایگزینی باز به خصوص در انتهای 3′ مشاهده میشود. خوانشهای جفت شده پایانی همراه با تولید رشتهای میتوانند بیان کنند که خطا رخ داده است یا خیر. رشتههای معکوس و پیشرو باید مکمل یکدیگر باشند، همه خوانشهای معکوس و خوانشهای رشته پیشرو باید با یکدیگر مطابقت داشته باشند. اگر رشته خوانش شده به اندازه کافی شبیه همتایان خود نباشد (شبیه سازی)، ممکن است خطایی رخ داده باشد. حداقل آستانه شباهت در آنالیزهای برخی آزمایشگاهها 97 درصد گزارش شده است.
توالی بر اساس سنتز:
در پایان تکثیر کلونال، تمام رشتههای معکوس از سلول هدف شسته شده و فقط رشتههای پیشرو باقی میمانند. یک پرایمر به محل اتصال پرایمر آداپتور رشتههای جلویی متصل میشود و یک پلیمراز، یک dNTP حاوی فلورسنت را به رشته DNA اضافه میکند. فقط یک dNTP حاوی فلورسنت میتواند در هر دور به DNA اضافه شود زیرا فلور به عنوان یک گروه مسدود کننده عمل میکند که حالت برگشت پذیری دارد. با استفاده از ویژگیهای شیمی چهار رنگ، هر یک از چهار رنگ پایه، انتشار منحصر به فردی دارند و پس از هر دور، دستگاه ضبط میکند که کدام قطعه اضافه شده است. پس از ثبت رنگ، فلور و dNTP غیرمنتظره شسته شده و فرآیند تکرار میشود.
با شروع NextSeq و بعداً MiniSeq، ایلومینا یک توالی دو رنگ جدید شیمیایی را معرفی کرد. نوکلئوتیدها با یکی از دو رنگ (قرمز یا سبز)، بدون رنگ (سیاه) یا ترکیب هر دو رنگ (به رنگ نارنجی به عنوان مخلوطی بین قرمز و سبز ظاهر میشوند) متمایز میشوند.
هنگامی که رشته DNA خوانده شد، رشتهای که به تازگی به رشته اضافه شده است، شسته میشود. سپس پرایمر شاخص 1 نشان دار شده، توالی شاخص 1 را پلیمریزه میکند و در نهایت شسته شده و حذف میشود. این رشته مجدداً یک پل را تشکیل میدهد و انتهای 3′ رشته DNA به الیگو سلول هدف میچسبد. پرایمر شاخص 2 نشان دار شده، نیز توالی شاخص 2 را پلیمریزه کرده و شسته میشود. پلیمراز، رشته مکمل را در بالای رشته قوسی قرار میدهد. رشته پیشرو شسته شده و روند سنتز توالی برای رشته معکوس تکرار میشود.
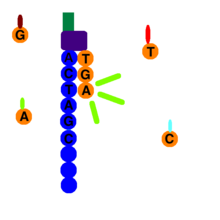
نوکلئوتیدهای برچسب زده شده به رشته DNA اضافه میشوند. هر یک از چهار نوکلئوتید دارای یک برچسب شناسایی هستند که میتواند برای انتشار یک طول موج مشخص برانگیخته شود. کامپیوتر همه نورهای منتشر شده را ثبت میکند.
تحلیل دادهها:
توالی یابی برای میلیونها رشته به طور همزمان انجام میشود و هر رشته دارای 1000 نسخه یکسان از یک نمونه DNA است. دادههای توالی بدست آمده با در نظر گرفتن مناطق همپوشانی، به نام contigs، و ردیف کردن آنها تجزیه و تحلیل میشوند. اگر مورد بررسی یک دنباله مرجع شناخته شده باشد، از contig ها برای شناسایی استفاده میشوند.
این فرآیند به دانشمندان اجازه میدهد تا توالی کامل را مشاهده کنند. با این حال، از آنجایی که طول خواندن Illumina خیلی طولانی نیست (توالی HiSeq میتواند خوانشی به طول حدود 90 جفت باز ایجاد کند)، میتواند روشی مناسب برای بررسی مناطق تکراری کوتاه tandem، باشد. همچنین، اگر دنباله de novo باشد و مرجعی وجود نداشته باشد، نواحی تکراری میتوانند ایجاد دنباله را با مشکل روبهرو کنند. دیگر مشکلات شامل جایگزینی قطعه (به ویژه در انتهای 3′ خوانده شده) توسط پلیمرازهای نادرست، توالیهای کایمریک و PCR-bias است که همگی میتوانند به ایجاد یک توالی نادرست منجر شوند.
مقایسه با سایر روش های توالی یابی:
این تکنیک نسبت به روش های توالی یابی قدیمی مانند توالی یابی سنگر مزیتهای متعددی دارد. تعیین توالی سنگر به دو واکنش نیاز دارد، یکی برای پرایمر فوروارد و دیگری برای پرایمر ریورس. بر خلاف ایلومینا، توالی یابی سنگر از تری فسفاتهای دی اکسی نوکلئوزیدی نشان دار شده با فلورسنت (ddNTPs) برای تعیین توالی قطعه DNA استفاده میکند. ddNTP ها گروه 3′ OH را ندارند و سنتز DNA را برای همیشه به پایان میرسانند. در هر لوله واکنش، dNTPs و ddNTPs به همراه DNA پلیمراز و پرایمرها اضافه میشوند. نسبت ddNTP ها به dNTP ها مهم است زیرا DNA الگو باید به طور کامل سنتز شود، و فراوانی بیش از حد ddNTP ها قطعات متعددی با اندازه و موقعیت یکسان الگوی DNA ایجاد میکند. هنگامی که DNA پلیمراز یک ddNTP اضافه میکند، سنتز قطعه پایان مییابد و یک قطعه جدید سنتز میشود. هر قطعه تولید شده به اندازه یک نوکلئوتید از آخرین رشته سنتز شده، طویلتر است. هنگامی که الگوی DNA به طور کامل سنتز شد، قطعات توسط الکتروفورز مویرگی جدا میشوند. در پایین لوله موئین، لیزر ddNTP های دارای فلورسنت را تحریک کرده و یک دوربین رنگ ساطع شده را دریافت میکند.
به دلیل ماهیت خودکار توالی یابی رنگ ایلومینا، میتوان چندین رشته را به طور همزمان توالی یابی کرد و دادههای توالی واقعی را به سرعت به دست آورد. با توالی یابی سنگر، تنها یک رشته میتواند در یک زمان توالی یابی شود و فرآیندی نسبتا کند است. توالی یابی ایلومینا تنها از DNA پلیمراز استفاده میکند که برخلاف آنزیمهای گران قیمت سایر تکنیکهای توالی یابی، مقرون به صرفه است.
نمونههایی از استفاده این روش توالی یابی:
توالی Illumina برای تحقیق در مورد رونوشتهای سیب زمینی شیرین و جنس ژیمنوسپرم Taxus استفاده شده است.
مترجم: امید آهنگریان ابهری
مطالعه صدها مطلب علمی در حوزه بیولوژی
آرشیو جدیدترین خبرهای روز دنیای بیولوژی
