


اطلاعات عمومی,ویکی ژن
قطعات اوکازاکی: تعریف،تشکیل، کاربرد و اهمیت آن
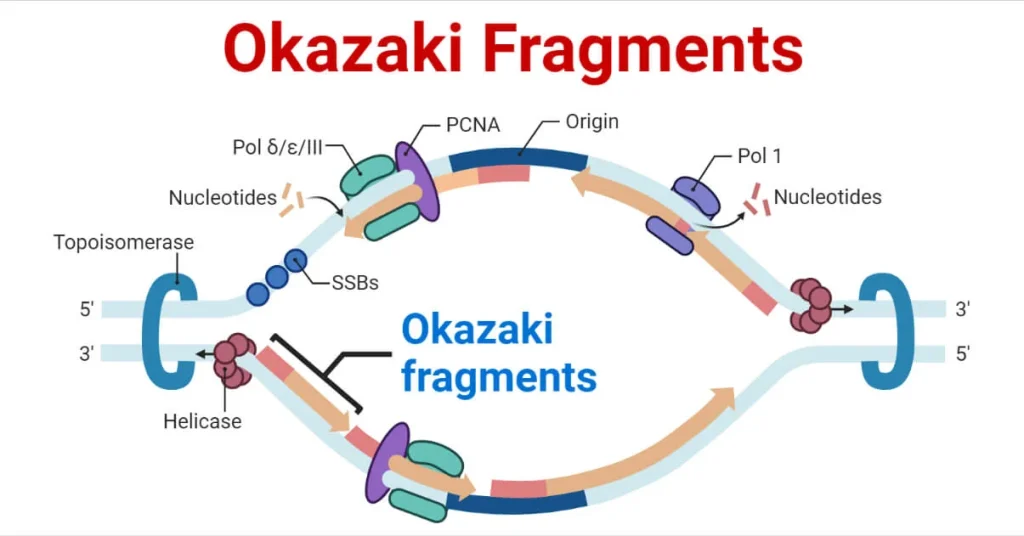
قطعات اوکازاکی چیست؟
قطعات اوکازاکی، نامگذاری شده به نام زیستشناسان مولکولی ژاپنی، رجی و تسونکو اوکازاکی، که آنها را در دهه ۱۹۶۰ شناسایی کردند، توالیهای کوتاه DNA هستند که در طول تکثیر رشته لگینگ (Lagging) DNA سنتز میشوند. این قطعات بهطور خاص روی رشتهای که جهت سنتز آن مخالف جهت حرکت چنگال تکثیر است، ساخته میشوند.
این قطعات اجزای ضروری در فرآیند تکثیر نیمهپیوسته هستند و اطمینان حاصل میکنند که کد ژنتیکی با دقت تکثیر میشود. فرآیند سنتز قطعات اوکازاکی به این صورت است که برای هر قطعه، یک پرایمر RNA ابتدا به رشته DNA متصل میشود تا نقطه شروعی برای پلیمریزاسیون DNA ایجاد کند. سپس آنزیم پلیمراز DNA این قطعات را از نو میسازد.
تکثیر DNA یک فرآیند پیچیده و بسیار تنظیمشده است که اطمینان حاصل میکند که اطلاعات ژنتیکی از یک نسل سلول به نسل بعدی به طور دقیق منتقل میشود. در طول این فرآیند، مولکول DNA دو رشتهای توسط آنزیم هلیکاز DNA باز میشود و ساختاری به نام چنگال تکثیر ایجاد میکند.
در این چنگال، سنتز رشتههای جدید DNA مکمل آغاز میشود که توسط آنزیمهایی مانند پرایماز DNA و پلیمراز DNA تسهیل میشود. [پرایماز، RNA پرایمر را برای شروع سنتز رشته جدید قرار میدهد و پلیمراز DNA با استفاده از این پرایمر، رشته مکمل جدید را سنتز میکند.]
در رشته لگینگ (Lagging)، پرایماز DNA پرایمرهای کوتاه RNA را سنتز میکند که به عنوان نقاط شروع برای پلیمراز DNA عمل میکنند. سپس پلیمراز این پرایمرها را گسترش میدهد و قطعات اوکازاکی را ایجاد میکند. این قطعات، در سلولهای یوکاریوتی، معمولاً بین ۱۰۰ تا ۲۰۰ نوکلئوتید طول دارند.
پس از سنتز، پرایمرهای RNA با DNA جایگزین میشوند و آنزیم لیگاز DNA شکافهای بین قطعات را مهر و موم میکند و در نتیجه یک رشته DNA پیوسته ایجاد میشود. {این فرآیند در راستای تکثیر نیمهپیوسته DNA است که هدف آن ایجاد دو رشته تکثیر شدهای از DNA است که هر دو، کد ژنتیکی دقیق را از مولکول والدین به نسلهای جدید منتقل کنند.}
کشف قطعات اوکازاکی یک نقطه عطف مهم در زیستشناسی مولکولی بود. از طریق آزمایشهای روی باکتری اشریشیا کلی، اوکازاکیها نشان دادند که تکثیر DNA در رشته لگینگ (Lagging) پیوسته نیست، همانطور که قبلاً تصور میشد. در عوض، آنها وجود قطعات کوتاه DNA را شناسایی کردند که بعدها قطعات اوکازاکی نامیده شدند، به عنوان واسطههای گذرا در فرآیند تکثیر.
{این کشف، بهویژه در زمینه پیریزی مفاهیم جدید در زمینه مکانیسمهای مولکولی، بهطور خاص با توجه به دینامیک دقیق عملکرد آنزیمها و نقش کلیدی آنها در فرآیند بازسازی و صحت تکثیر DNA، اهمیت ویژهای پیدا کرد.}
در خلاصه، قطعات اوکازاکی در فرآیند تکثیر DNA بسیار مهم هستند و اطمینان حاصل میکنند که رشته لگینگ (Lagging) با دقت و کامل تکثیر میشود. کشف آنها درک ما از سنتز DNA را تغییر داد و پیچیدگیها و دقت فرآیندهای سلولی را برجسته کرد.
{این کشف نهتنها به پردازش بهتر فرآیند تکثیر DNA کمک کرده است، بلکه در مطالعات ژنتیکی و سلولی نیز راههای جدیدی برای درک بیماریها و اختلالات ژنتیکی فراهم آورده است.}
قطعات اوکازاکی چیستند؟
قطعات اوکازاکی توالیهای کوتاه DNA هستند که به صورت گسسته در رشته لگینگ (Lagging) در طول تکثیر DNA سنتز میشوند و بعدا به هم متصل میشوند تا یک رشته پیوسته تشکیل دهند.
چرا قطعات اوکازاکی تشکیل میشوند؟
قطعات اوکازاکی به دلیل ماهیت ذاتی پلیمراز DNA و ساختار ضد موازی DNA تشکیل میشوند. در اینجا توضیحی در مورد اینکه چرا قطعات اوکازاکی تشکیل میشوند، ارائه شده است:
۱. ساختار ضد موازی DNA:
- DNA از دو رشته تشکیل شده است که در جهت مخالف یکدیگر قرار دارند: یکی از ۵’ به ۳’ و دیگری از ۳’ به ۵’. اینها اغلب به عنوان رشتههای ضد موازی مارپیچ مضاعف DNA نامیده میشوند. {این ویژگی ساختاری موجب پیچیدگیهایی در تکثیر DNA میشود زیرا آنزیمها برای تکثیر DNA باید با جهتگیری متفاوت رشتهها تطابق یابند.}
۲. جهتگیری پلیمراز DNA:
- پلیمراز DNA، آنزیمی که مسئول سنتز رشتههای جدید DNA است، فقط میتواند نوکلئوتیدها را به انتهای ۳’ یک رشته DNA در حال رشد اضافه کند. این بدان معناست که پلیمراز DNA فقط میتواند DNA را در جهت ۵’ به ۳’ سنتز کند.
- بنابراین، در حالی که یکی از رشتهها به طور پیوسته و پیوسته در جهت ۵’ به ۳’ تکثیر میشود، رشته دیگر به دلیل جهت مخالف، نمیتواند به صورت پیوسته تکثیر شود.
۳. سنتز پیوسته رشته لیدینگ:
- هنگامی که مارپیچ مضاعف DNA برای تکثیر باز میشود، یکی از رشتهها (رشته لیدینگ) به گونهای جهتگیری شده است که انتهای ۳’ آن به سمت چنگال تکثیر قرار دارد. این به پلیمراز DNA اجازه میدهد تا رشته لیدینگ (Leading) را به طور پیوسته در جهت ۵’ به ۳’ سنتز کند زیرا چنگال پیشرفت میکند.
- سنتز پیوسته در رشته لیدینگ باعث میشود که این رشته به طور همزمان با پیشروی چنگال تکثیر، در همان جهت سنتز شود. این فرآیند به طور مداوم تکرار میشود و اجازه میدهد که فرآیند تکثیر بدون توقف ادامه یابد.
۴. سنتز گسسته رشته لگینگ (Lagging):
- رشته دیگر، که به آن رشته لگینگ (Lagging) گفته میشود، به گونهای جهتگیری شده است که انتهای ۳’ آن در خلاف جهت چنگال تکثیر قرار دارد. این وضعیت باعث میشود که پلیمراز DNA قادر به انجام سنتز پیوسته در این رشته نباشد، چرا که آنزیم پلیمراز تنها توانایی افزودن نوکلئوتیدها به انتهای ۳’ یک رشته در حال رشد را دارد. بدین ترتیب، با پیشرفت چنگال تکثیر و در دسترس قرار گرفتن بخشهای جدید از الگوی رشته لگینگ (Lagging)، سنتز DNA به صورت گسسته و به فاصلههای کوتاه انجام میشود. این سنتز گسسته موجب تشکیل قطعات کوتاه و مجزای DNA میشود که به آنها قطعات اوکازاکی گفته میشود.
- این فرآیند اساساً نتیجه ماهیت ضد موازی ساختار دو رشتهای DNA است که الزاماً ایجاب میکند برای جبران جهتگیری متناقض دو رشته، یک بخش از الگوی DNA به صورت گسسته و ناپیوسته سنتز شود. این سنتز گسسته در واقع پاسخ بیولوژیکی به چالشهای ساختاری است که در تکثیر رشته ضد موازی باید رفع شود.
۵. نقش پرایمرهای RNA:
- سنتز هر قطعه اوکازاکی با قرار گرفتن یک پرایمر RNA آغاز میشود. این پرایمر که از حدود ۱۰ تا ۱۵ نوکلئوتید تشکیل شده است، به انتهای ۳’ برای شروع سنتز DNA متصل میشود. این پرایمر، با فراهم کردن یک انتهای ۳’ هیدروکسیل برای آنزیم پلیمراز DNA، شروع به افزودن نوکلئوتیدها میکند.
- به محض ادامه پیشروی چنگال تکثیر و نمایان شدن بخشهای جدید از رشته لگینگ (Lagging)، یک پرایمر RNA جدید بر روی الگوی DNA قرار میگیرد و سنتز قطعه جدید اوکازاکی آغاز میشود. {در این مرحله، پرایمر RNA به عنوان یک عامل ضروری عمل میکند، زیرا پلیمراز DNA نمیتواند بدون وجود یک انتهای ۳’ هیدروکسیل (OH) فعالیت کند. بنابراین، عملکرد پرایمر RNA نه تنها بهعنوان نقطه آغاز، بلکه به عنوان یک پیشنیاز اساسی در سنتز DNA با دقت بالا میباشد.}
۶. تکمیل رشته لگینگ (Lagging):
- پس از سنتز هر قطعه اوکازاکی، پرایمرهای RNA به سرعت از قطعات جدا شده و با نوکلئوتیدهای DNA جایگزین میشوند. در مرحله بعد، آنزیم لیگاز DNA، با ایجاد پیوند فسفودیاستری بین نوکلئوتیدهای مجاور، شکافهای باقیمانده بین قطعات اوکازاکی را مهر و موم کرده و به این ترتیب یک رشته پیوسته و کامل از DNA را تشکیل میدهد.
- این فرآیند مهر و موم کردن شکافها و تکمیل سنتز رشته لگینگ (Lagging) موجب میشود که رشته لگینگ (Lagging) نیز بهطور مشابه با رشته لیدینگ (Leading) بهطور دقیق و بدون هیچ گونه اختلالی تکثیر شود. {این مرحله، که در آن آنزیم لیگاز بهطور دقیق شکافهای میان قطعات اوکازاکی را بازسازی میکند، از نظر فرآیندی حائز اهمیت است. چرا که این عمل، که مستلزم برقراری پیوندهای کووالانسی در ساختار دو رشتهای DNA است، منجر به ایجاد یک مولکول DNA واحد و پیوسته میشود.}
به طور خلاصه، قطعات اوکازاکی در رشته لگینگ (Lagging) در طول تکثیر DNA به دلیل ماهیت ضد موازی ساختار DNA و جهتگیری منحصر به فرد آنزیم پلیمراز DNA در فرآیند سنتز تشکیل میشوند. این فرآیند سنتز گسسته بهطور دقیق از طریق سیستمهای مولکولی مختلف تنظیم میشود که هرکدام از آنها بهطور ویژه برای رفع چالشهای ساختاری و جهتگیریهای متناقض دو رشته DNA طراحی شدهاند.
از اینرو، تشکیل قطعات اوکازاکی نه تنها یک راهحل بیولوژیکی پیچیده برای تکثیر دقیق و بدون نقص DNA است، بلکه نمایانگر پیچیدگیهای زیستی است که در سطح مولکولی در سیستمهای سلولی برای حفظ یکپارچگی اطلاعات ژنتیکی بهکار گرفته میشود.
کشف قطعات اوکازاکی
- کشف قطعات اوکازاکی نقطه عطفی در درک تکثیر DNA بود. در اوایل دهه ۱۹۶۰، تسونکو اوکازاکی و رئیجی اوکازاکی، محققان دانشگاه ناگویا در ژاپن، تحقیقی را برای روشنسازی مکانیسمهای زیرین تکثیر DNA آغاز کردند. فرضیه آنها بر این ایده متمرکز بود که تکثیر نیمهمحافظهکارانه DNA ممکن است شامل یک مکانیزم سنتز غیرپیوسته برای رشتههای دختر باشد.
- این تحقیق بهطور خاص به بررسی رفتار رشته لگینگ (Lagging) در فرآیند تکثیر DNA میپرداخت و اهمیت آن به این دلیل بود که تا آن زمان تصور میشد که هر دو رشته DNA به صورت پیوسته و به طور همزمان سنتز میشوند.
- اوکازاکیها پیشنهاد کردند که رشته لگینگ (lagging strand) که در جهت ۳′ به ۵′ رشد میکند، ممکن است به صورت قطعات کوتاه در جهت ۵′ به ۳′ سنتز شود که مخالف جهت گسترش واقعی آن است. با اتصال این قطعات به یکدیگر، رشته دختر میتواند طولانی شود.
- این ایده، که به عنوان مدل سنتز گسسته شناخته میشود، برخلاف پیشفرضهای قبلی بود که معتقد بودند هر دو رشته DNA به طور پیوسته سنتز میشوند. در واقع، این مدل گویای پیچیدگیهای ساختاری و مکانیکی فرآیند تکثیر DNA بود که به دلیل ساختار ضد موازی آن و محدودیتهای آنزیمهای سنتز DNA، نیازمند راهحلهایی بود که امکان رشد هر دو رشته را فراهم کند.
- برای تایید این فرضیه، آنها از آزمایش پالس-چیس (pulse-chase) استفاده کردند. آنها به طور موقت DNA که به طور فعال در حال تکثیر بود را در معرض نوکلئوتیدهای تریتیهشده (tritiated nucleotides) قرار دادند، به طوری که این نوکلئوتیدهای “داغ” وارد رشتههای DNA شدند. پس از این پالس، DNA به سرعت استخراج شده و برای مدتهای مختلف در معرض نوکلئوتیدهای “سرد” بدون برچسب قرار گرفت.
- آزمایش پالس-چیس یک روش هوشمندانه بود که امکان ردیابی تکثیر DNA در مقیاس زمانی محدود و با دقت بالا را فراهم میکرد. استفاده از نوکلئوتیدهای رادیواکتیو این امکان را به محققان میداد که به طور دقیق زمانبندی فرآیند سنتز و نحوه پیوستن قطعات DNA را بررسی کنند.
- سانتریفیوژ نمونههای DNA نشان داد که پس از مدت زمان کوتاه پالس، بیشتر رادیواکتیویته با قطعات کوچک DNA مرتبط بود. با این حال، با افزایش مدت زمان پالس، رادیواکتیویته در رشتههای DNA بزرگتر یافت شد، که نشاندهنده این بود که قطعات کوچک به مرور زمان به رشتههای بزرگتر پیوستهاند.
- این مشاهده تأیید کرد که قطعات کوتاه DNA (قطعات اوکازاکی) در طول تکثیر رشته لگینگ (Lagging) به طور موقت وجود دارند و سپس با تکمیل فرآیند، به یک رشته پیوسته تبدیل میشوند. این یافته نقطه عطفی در درک فرآیند سنتز DNA بود، زیرا نشان داد که تکثیر رشته لگینگ (Lagging) برخلاف آنچه پیشتر تصور میشد، یک فرآیند گسسته است.
- این آزمایش انقلابی شواهدی برای وجود قطعات کوتاه DNA در حین سنتز رشته لگینگ فراهم کرد. این قطعات بعدها “قطعات اوکازاکی” نامگذاری شدند تا یادبود کشفکنندگان آنها باشد، این نامگذاری در سال ۱۹۶۸ توسط رولین هاتچکیش (Rollin Hotchkiss) در همایش Cold Spring Harbor پیشنهاد شد.
- این نامگذاری نه تنها یادآور تلاشهای علمی تسونکو و رئیجی اوکازاکی بود، بلکه به عنوان یکی از مفاهیم اساسی در زیستشناسی مولکولی شناخته شد که بهطور شگرفی درک ما از پیچیدگیهای فرآیند تکثیر DNA را بهبود بخشید.
- آزمایشهای پشتیبانیکننده بیشتر شامل برچسبگذاری پالس (pulse-labeling) (Escherichia coli) با (3H-thymidine)
³H-thymidine تحت شرایطی بود که نرخ رشد و تقسیم آنها را کاهش میداد. باکتریها ابتدا در دمای ۳۷ درجه سلسیوس با ¹⁴C-thymidine کشت داده شدند تا DNA آنها به طور یکنواخت برچسبگذاری شود. - این روش برای اطمینان از اینکه همه مولکولهای DNA در باکتریها به صورت یکسان برچسبگذاری شدهاند و نتایج آزمایش دقیق و قابل تکرار باشد، انجام شد.
- پس از آن، دما به ۲۰ درجه سلسیوس کاهش داده شد و سلولها به مدت ۱۰ ثانیه با ³H-thymidine (3H-thymidine) برچسبگذاری شدند. این روش هدف داشت تا واسطههای گذرا در تکثیر DNA تحت شرایط کاهش یافته تکثیر شناسایی شوند.
- کاهش دما به منظور کند کردن فرآیند تکثیر و کاهش سرعت سنتز DNA انجام شد، به طوری که قطعات کوتاهتر از DNA به طور موقت شکل بگیرند و امکان شناسایی این واسطهها را فراهم آورد. این استراتژی در واقع به دانشمندان کمک کرد که فرآیندهایی را که در حال وقوع هستند، در زمانهای خاص ردیابی کنند.
- پس از پالس، DNA استخراج شده و تحت سانتریفیوژ در گرادیان ساکروز قلیایی قرار گرفت. تجزیه و تحلیل فراکشنهای گرادیان نشان داد که بیشتر DNA برچسبگذاریشده با ³H (3H-thymidine) در ابتدا به صورت قطعاتی با طول ۵۰ تا ۵۰۰۰ نوکلئوتید ظاهر شد. با گذشت زمان، این قطعات طولانی شده و ادغام شدند، که به عنوان واسطههای گذرا در تکثیر DNA عمل کردند.
- این مشاهدات به وضوح نشان داد که قطعات کوتاه DNA که در ابتدا به صورت قطعات گسسته ظاهر میشوند، به تدریج به یک ساختار پیوستهتر تبدیل میشوند. این نشاندهنده فرآیند تشکیل قطعات اوکازاکی و پیوستن آنها به یکدیگر در طی تکثیر رشته لگینگ (Lagging) بود.
- در نتیجه، آزمایشهای دقیق انجام شده توسط اوکازاکیها وجود قطعات کوتاه DNA، که امروزه به نام قطعات اوکازاکی شناخته میشوند، را آشکار کرد که نقش حیاتی در سنتز غیرپیوسته رشته لگینگ در طول تکثیر DNA ایفا میکنند. {این کشف یکی از دستاوردهای مهم در زیستشناسی مولکولی بود که پایهگذار درک عمیقتری از مکانیزمهای پیچیده تکثیر DNA گردید. کشف قطعات اوکازاکی نقش کلیدی در پیشبرد فهم فرآیند تکثیر نیمهمحافظهکارانه و اطمینان از دقت آن داشت.}
چرا قطعات اوکازاکی ناپیوسته هستند؟
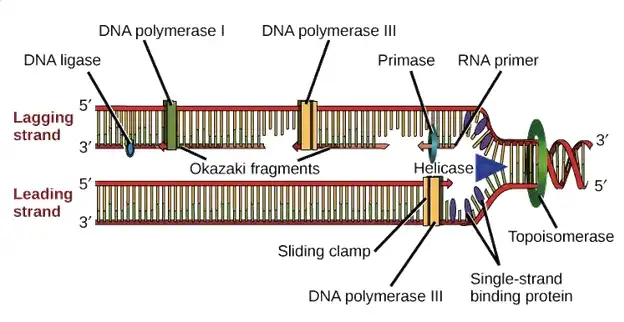
- قطعات اوکازاکی، توالیهای کوتاه DNA هستند که در فرآیند تکثیر DNA ساخته میشوند. کشف و درک این قطعات به کار پیشگامانه کیواکو ساکابه (Kiwako Sakabe)، رئیجی اوکازاکی (Reiji Okazaki)، و تسونکو اوکازاکی (Tsuneko Okazaki) نسبت داده میشود. در گذشته، باور غالب در جامعه علمی این بود که تکثیر DNA به طور پیوسته در هر دو جهت ۳′ به ۵′ و ۵′ به ۳′ انجام میشود.
- اعداد ۳′ و ۵′ به کربنهای خاصی در حلقه دئوکسی ریبوز در اسیدهای نوکلئیک اشاره دارند که جهتگیری یا جهتدهی رشته DNA را نشان میدهند. {این تصور غالب که تکثیر DNA به صورت پیوسته و در هر دو جهت انجام میشود، بر اساس مشاهدات اولیه از مکانیزمهای تکثیر در برخی از ارگانیسمها بود، اما آزمایشات بعدی پیچیدگیهای بیشتری را آشکار کرد.}
- در سال ۱۹۶۷، یک پیشنهاد انقلابی توسط تسونکو اوکازاکی و تورو اوگاوا (Toru Ogawa) مطرح شد. آنها اظهار داشتند که هیچ مکانیزم شناخته شدهای اجازه تکثیر پیوسته در جهت ۳′ به ۵′ را نمیدهد. بلکه تکثیر در این جهت ممکن است به صورت غیرپیوسته باشد، به طوری که DNA به صورت قطعات کوتاه سنتز شود.
- این قطعات سپس در جهت ۵′ به ۳′ به رشته پیشموجود متصل میشوند، که این فرآیند با کمک آنزیم DNA پلیمراز (DNA polymerase) تسهیل میشود. {این فرضیه، که پایهگذار مفهوم تکثیر غیرپیوسته در رشته لگینگ است، با مفهوم سنتز پیوسته در رشته پیشموجود در تضاد است و به یکی از کشفیات کلیدی در زیستشناسی مولکولی تبدیل شد.}
- برای تأیید این فرضیه، تیم تحقیقاتی از یک روش تجربی استفاده کردند که در آن نواحی جدیداً تکثیر یافته کروموزومهای (Escherichia coli) با رادیواکتیو نشانهگذاری شدند. با دناتوره کردن و استخراج DNA، آنها حضور قابل توجهی از واحدهای کوتاه DNA رادیواکتیو را مشاهده کردند که نشاندهنده مکانیزم تکثیر غیرپیوسته بود. این فرضیه با شناسایی لیگاز پلینوکلئوتیدی (polynucleotide ligase)، آنزیمی که مسئول اتصال این رشتههای کوتاه DNA است، قوت بیشتری گرفت.
- این شواهد تجربی نه تنها فرضیه سنتز قطعات کوتاه DNA را تایید کرد بلکه نشان داد که پیوستن این قطعات به یکدیگر، به کمک آنزیم لیگاز، نقش تعیینکنندهای در تکمیل فرآیند تکثیر داشت.
- در آزمایش بعدی در سال ۱۹۶۸، اوکازاکیها شواهد بیشتری ارائه دادند. آنها اظهار داشتند که اگر سنتز DNA واقعاً شامل تکثیر غیرپیوسته باشد، آنگاه شرایطی که عملکرد لیگاز را مختل کند، باید منجر به انباشته شدن این زنجیرههای کوتاه DNA شود. با استفاده از (Escherichia coli) آلوده به باکتریوفاژ خاص T4 که لیگاز پلینوکلئوتیدی حساس به دما تولید میکند، آنها مشاهده کردند که در دماهای بالاتر، انباشت قابل توجهی از زنجیرههای کوتاه DNA مشاهده میشود. این مشاهده نه تنها فرضیه آنها را تقویت کرد بلکه ایدهای را که این زنجیرههای کوتاه فقط آثار جانبی فرآیند استخراج بودند، رد کرد.
- این آزمایشها، به ویژه آزمایشهایی که شرایط دما را تغییر میدادند، نشان داد که تکثیر غیرپیوسته در واقع یک جزء اصلی از فرآیند تکثیر DNA است و نه فقط یک پدیده غیرعمدی.
- به طور خلاصه، آزمایشهای دقیق انجام شده توسط اوکازاکیها ماهیت غیرپیوسته تکثیر DNA، بهویژه در رشته لگینگ (lagging strand)، را روشن ساخت. این منجر به شناسایی و درک آنچه که اکنون به عنوان قطعات اوکازاکی (Okazaki fragments) شناخته میشود، گردید. این قطعات نقش کلیدی در تضمین تکثیر دقیق DNA ایفا میکنند و پیچیدگیها و دقت موجود در فرآیندهای سلولی را برجسته میسازند.
- این کشف که تکثیر در رشته لگینگ به صورت گسسته و از طریق تشکیل قطعات کوتاه انجام میشود، به درک ما از دقت و کارآمدی فرآیند تکثیر DNA افزوده و تاثیرات عمیقی در زیستشناسی مولکولی داشت.
تشکیل قطعات اوکازاکی – چگونه قطعات اوکازاکی سنتز میشوند؟
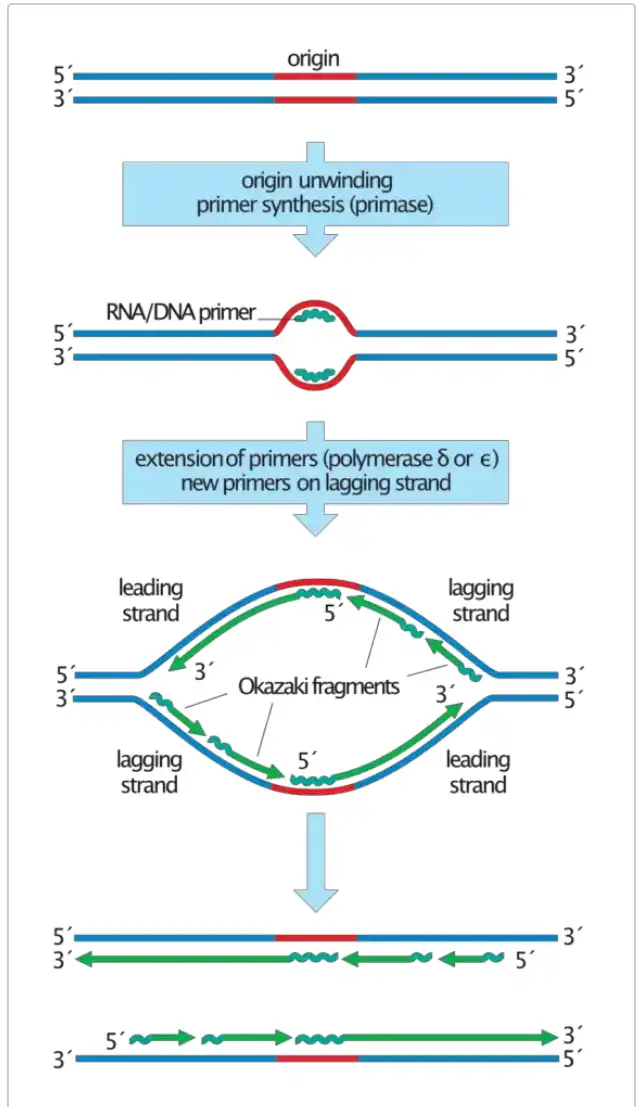
- تشکیل قطعات اوکازاکی یک جنبه اساسی از تکثیر DNA است، به ویژه در مورد سنتز رشته لگینگ (lagging strand). این فرآیند با تشکیل فورک تکثیر DNA آغاز میشود، که ساختاری است که هنگام باز شدن دوکلافه (double helix) DNA به وجود میآید. این باز شدن توسط آنزیم هلیکاز (DNA helicase) تسهیل میشود که رشتههای مکمل DNA را از یکدیگر جدا میکند.
- {این باز شدن و جداسازی رشتههای DNA باعث ایجاد دو رشته جداگانه میشود که به عنوان قالب برای سنتز رشتههای جدید عمل میکنند. حرکت این فورک تکثیر به صورت پیوسته و هماهنگ با دیگر آنزیمهای درگیر در فرآیند تکثیر انجام میشود.}
- با پیشرفت تکثیر، آنزیمهایی مانند DNA پریماز (DNA primase) و DNA پلیمراز (DNA polymerase) نقشهای مهمی در سنتز رشته مکمل جدید ایفا میکنند. با این حال، جهتگیری ذاتی این آنزیمها که تنها در جهت ۵′ به ۳′ عمل میکنند، چالشی ایجاد میکند. برای رشته پیشرو (leading strand)، که قالبی در جهت ۳′ به ۵′ دارد، تکثیر به صورت پیوسته است و DNA پلیمراز رشته را به طور همزمان با پیشرفت فورک تکثیر سنتز میکند.
- {در این راستا، تکثیر رشته پیشرو بدون وقفه و همزمان با حرکت آنزیمها در جهت حرکت فورک تکثیر صورت میگیرد. این پیوستگی در سنتز باعث کاهش خطاهای تکثیر میشود.}
- برعکس، رشته لگینگ که جهت قالب آن ۵′ به ۳′ است، نمیتواند به طور پیوسته سنتز شود. بلکه تشکیل آن با وقفههای مکرر همراه است. سنتز رشته لگینگ با شروع از RNA پرایمرها، که توسط پریماز تولید میشوند، آغاز میشود.
- {این وقفهها به دلیل جهتگیری مخالف آنزیمها با جهت حرکت فورک تکثیر ایجاد میشود، که منجر به ساخت قطعات کوچک و گسسته از DNA میگردد.}
- این پرایمرها به عنوان نقاط شروع برای DNA پلیمراز δ در یوکاریوتها (و DNA پلیمراز I در پروکاریوتها) عمل میکنند تا رشته را گسترش دهند. به دلیل حرکت متقابل پریماز و پلیمراز نسبت به فورک تکثیر، این آنزیمها باید به طور مکرر متوقف شده و دوباره شروع به کار کنند، فرآیندی که توسط DNA هلیکاز تسهیل میشود.
- {این حرکت دینامیک آنزیمها، که در آن به طور مداوم متوقف و شروع به کار میکنند، به تکثیر دقیق رشته لگینگ کمک میکند.}
- برای سنتز هر قطعه اوکازاکی، به یک RNA پرایمر جدید نیاز است. فرآیند پرایمرسازی به صورت تدریجی انجام میشود: ابتدا پروتئین PriA پروتئینهای SSB را از یک بخش DNA جابجا میکند، سپس پریماز (DnaG) به PriA متصل میشود و در نهایت، پریماز یک RNA پرایمر ۱۱-۱۲ بازهای را سنتز میکند.
- {این فرآیند تدریجی و هماهنگ از یکپارچگی قطعات و جلوگیری از تداخل آنزیمها اطمینان میدهد.}
- با پیشرفت سنتز رشته لگینگ، DNA پلیمراز چنگ خود را از روی DNA رها کرده، به انتهای ۳′ یک RNA پرایمر جدید منتقل شده و سنتز یک بخش جدید DNA آغاز میشود. این حرکت دینامیک توسط کمپلکس بارگذاری کلمپ (clamp-loading complex) مدیریت میشود که مسئول تجزیه و انتقال کلمپ لغزنده است.
- {کمپلکس کلمپ به عنوان یک ساختار قابلانعطاف، تعامل آنزیمها را در طول فرآیند تکثیر هماهنگ میکند و باعث افزایش کارایی فرآیند میشود.}
- جالب اینجاست که تنها یکی از دو آنزیم هستهای DNA پلیمراز III در ریپلیزوم (replisome) این جابجایی را انجام میدهد، که اطمینان میدهد فقط رشته لگینگ تغییرات مکرر در موقعیت کلمپ را تجربه میکند.
- {این ویژگی به طور خاص به روند تکثیر رشته لگینگ محدود میشود و در رشته پیشرو، این تغییرات مکرر در موقعیت کلمپ رخ نمیدهد.}
- پس از سنتز، RNA پرایمرها باید حذف شوند تا اتصال قطعات اوکازاکی به صورت یکپارچه انجام شود. آنزیمهایی با فعالیت اندونوکلئولیتیک، مانند ریبونوکلئاز H (RNAse H)، فلاپ اندونوکلئازها (FENs)، و Dna2 هلیکاز/نوکلیازها این وظیفه را انجام میدهند
- {این فرآیند حذف و اصلاح، موجب پیوستگی کامل قطعات اوکازاکی میشود که در ادامه تکثیر DNA به صورت پیوسته و بدون شکستگی ادامه مییابد.}
- در پروکاریوتها، عملکرد نوکلیاز FEN در DNA پلیمراز I جای گرفته است، در حالی که در یوکاریوتها، FEN ها موجوداتی مجزا هستند. مکانیزم دقیق حذف RNA-DNA پرایمرها از قطعات اوکازاکی هنوز یکی از حوزههای تحقیقاتی فعال است.
- {پیشرفتهای علمی در درک نحوه و مکانیزم دقیق این اصلاحات به بهبود مدلهای موجود در تکثیر DNA کمک کرده است.}
- پس از حذف پرایمرها، DNA لیگاز (DNA ligase) قطعات را با استفاده از پیوندهای فسفودی استر به هم متصل میکند که در نتیجه آن یک رشته DNA پیوسته شکل میگیرد. این روش دوگانه تکثیر، که در آن یک رشته به طور پیوسته و دیگری به طور غیرپیوسته سنتز میشود، به عنوان تکثیر نیمهغیرپیوسته (semi-discontinuous replication) شناخته میشود.
- {این تکثیر نیمهغیرپیوسته، که در آن یکی از رشتهها به طور پیوسته و دیگری به صورت گسسته و غیرپیوسته تکثیر میشود، یکی از ویژگیهای بارز در فرآیند تکثیر DNA است.}
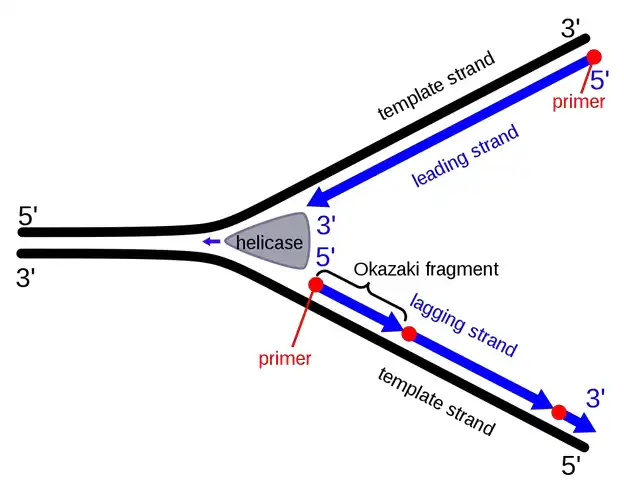
تشکیل قطعات اوکازاکی: فرآیند گام به گام
-
آغاز فورک تکثیر:
- دوکلافه DNA باز میشود.
- آنزیم DNA هلیکاز (DNA helicase) رشتههای مکمل DNA را از هم جدا کرده و فورک تکثیر تشکیل میشود.
-
فعالسازی آنزیمها:
- آنزیمهای DNA پریماز (DNA primase) و DNA پلیمراز (DNA polymerase) در فورک تکثیر فعال شده و سنتز رشته جدید مکمل DNA آغاز میشود.
-
سنتز رشته لیدینگ (Leading):
- رشته پیشرو با جهتگیری قالب 3′ به 5′، تکثیر پیوستهای را توسط DNA پلیمراز تجربه میکند که به طور هماهنگ با فورک تکثیر پیش میرود.
-
سنتز رشته لگینگ:
- به دلیل جهتگیری قالب 5′ به 3′ رشته لگینگ، DNA پلیمراز در جهت مخالف فورک تکثیر حرکت میکند.
- این حرکت منجر به وقفههای دورهای در فرآیند سنتز میشود.
-
آغاز قطعات اوکازاکی با RNA پرایمرها:
- DNA پریماز یک RNA پرایمر به عنوان نقطه شروع برای هر قطعه اوکازاکی سنتز میکند.
- DNA پلیمراز δ (در یوکاریوتها) یا DNA پلیمراز I (در پروکاریوتها) این پرایمرها را به سمت فورک تکثیر گسترش میدهند.
-
سنتز پرایمرهای جدید RNA:
- پروتئین PriA پروتئینهای SSB را از یک بخش از DNA جابجا میکند.
- پریماز (DnaG) به PriA متصل میشود.
- پریماز یک RNA پرایمر متشکل از 11-12 باز سنتز میکند.
-
جابجایی DNA پلیمراز:
- DNA پلیمراز در حال کار بر روی رشته لگینگ موقعیت فعلی خود را رها میکند.
- سپس به انتهای 3′ RNA پرایمر جدید منتقل شده و سنتز قطعه جدید اوکازاکی آغاز میشود.
-
دینامیک کلمپ لغزنده:
- کمپلکس بارگذاری کلمپ (clamp-loading complex) مسئول تجزیه و انتقال کلمپ لغزنده است.
- تنها یکی از دو آنزیم هستهای DNA پلیمراز III در ریپلیزوم این جابجایی را انجام میدهد که اطمینان میدهد فقط رشته لگینگ تغییرات مکرر در موقعیت کلمپ را تجربه میکند.
-
حذف RNA پرایمرها:
- آنزیمهایی با فعالیت اندونوکلئولیتیک مانند ریبونوکلئاز (RNAse H) H ، فلاپ اندونوکلئازها (FENs)، و Dna2 هلیکاز/نوکلیازها RNA پرایمرها را حذف میکنند.
- در پروکاریوتها، عملکرد نوکلیاز FEN توسط یک دامنه از DNA پلیمراز I انجام میشود.
-
لایگیشن قطعات اوکازاکی:
- هنگامی که قطعات اوکازاکی سنتز شده و RNA پرایمرها حذف شدند، DNA لیگاز (DNA ligase) قطعات را با استفاده از پیوندهای فسفودی استر به هم متصل میکند.
- این عمل منجر به تشکیل یک رشته DNA پیوسته میشود.
-
11. اتمام تکثیر:
- با توجه به سنتز پیوسته رشته پیشرو و سنتز غیرپیوسته رشته لگینگ، فرآیند کلی تکثیر DNA به عنوان تکثیر نیمهغیرپیوسته (semi-discontinuous replication) شناخته میشود.
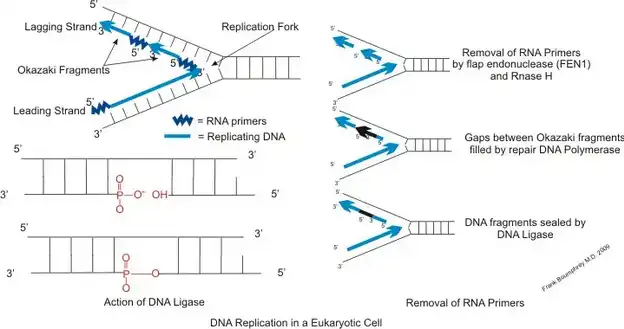
مسیرهای پردازش قطعات اوکازاکی
فرآیند پیچیده تکثیر DNA، به ویژه در سنتز رشته لگینگ، شامل تشکیل قطعات اوکازاکی است. پردازش صحیح این قطعات برای استمرار و یکپارچگی رشته جدید سنتز شده DNA بسیار مهم است. سه مسیر مختلف برای مدیریت پردازش این قطعات شناسایی شده است:
-
مسیر فلاپ کوتاه:
در تکثیر DNA یوکاریوتی، رشته لگینگ به طور موقت پرایمرگذاری میشود که منجر به تشکیل قطعات اوکازاکی میشود. مسیر فلاپ کوتاه عمدتاً شامل نوکلیاز FEN1 است. همانطور که DNA پلیمراز دلتا (Pol δ) رشته لگینگ را سنتز میکند، گاهی اوقات RNA/DNA پرایمر آغازگر را جابجا کرده و یک فلاپ 5′ تشکیل میشود. FEN1، که یک اندونوکلیاز 5′ به 3′ است، این فلاپ را شناسایی و برش میدهد تا برای لایگیشن بعدی آماده شود.
این مسیر اطمینان میدهد که پرایمر سنتز شده توسط Pol α حذف شود. فرآیند برش توسط مکانیزم ردیابی FEN1 تسهیل میشود که از فلاپ 5′ به سمت پایه آن حرکت میکند. سپس DNA لیگاز شکاف را میبندد و رشته DNA پیوسته تشکیل میشود. پروتئین PCNA (proliferating cell nuclear antigen) در این فرآیند نقش مهمی دارد و عملکردهای آنزیمی FEN1 و DNA لیگاز را تقویت میکند، به طوری که لایگیشن صحیح رشته لگینگ تضمین میشود.
-
مسیر فلاپ بلند:
در برخی موارد، FEN1 از کمپلکس ریپلیزوم به طور موقت جدا میشود که منجر به تشکیل فلاپهای بلند توسط Pol δ میشود. این فلاپهای بلند زمانی که توسط پروتئین(Replication Protein A) RPA بسته میشوند، نیاز به دخالت یک نوکلیاز جایگزین، یعنی DNA2 دارند.
DNA2 با FEN1 برای پردازش این فلاپهای بلند همکاری میکند. این آنزیم میتواند RPA را از فلاپ جدا کرده و با استفاده از مکانیزم مشابه FEN1، فلاپ را برش دهد و آن را برای برش توسط FEN1 و لایگیشن بعدی آماده کند. این مسیر که به آن “مسیر فلاپ بلند” گفته میشود، قابلیتهای مختلف دستگاه سلولی را در مدیریت چالشهای مختلف تکثیر نشان میدهد.
-
مسیر جایگزین:
تحقیقات اخیر یک مسیر اضافی برای پردازش قطعات اوکازاکی را آشکار کرده است. این مکانیزم جایگزین شامل عملکرد مشترک Pol δ و Pif1 است که به طور مشترک فرآیند حذف فلاپ را انجام میدهند، مشابه عملکرد مشترک Pol δ و FEN1. این کشف پیچیدگی و سازگاری فرآیند تکثیر DNA را نشان میدهد که اطمینان حاصل میکند اطلاعات ژنتیکی به طور صحیح و مداوم از نسلی به نسل دیگر منتقل میشود.
نتیجهگیری
پردازش صحیح قطعات اوکازاکی برای یکپارچگی و صحت تکثیر DNA ضروری است. این سه مسیر مختلف که شامل مسیر فلاپ کوتاه، مسیر فلاپ بلند و مسیر جایگزین هستند، فرآیندهای پیچیدهای را برای حذف پرایمرها و اتصال قطعات DNA به یکدیگر فراهم میآورند. این مکانیزمها نشاندهنده پیچیدگی و دقت بالای دستگاه سلولی در نگهداری یکپارچگی ژنتیکی در طول تکثیر هستند.
عملکرد قطعات اوکازاکی
قطعات اوکازاکی نقش بسیار مهمی در فرآیند پیچیده تکثیر DNA دارند و به طور مستقیم باعث اطمینان از تکثیر صحیح و کامل اطلاعات ژنتیکی میشوند. این فرآیند برای رشد و تقسیم سلولی ضروری است و اطمینان میدهد که اطلاعات ژنتیکی به درستی از یک نسل به نسل بعد منتقل میشود. در اینجا عملکرد قطعات اوکازاکی در تکثیر DNA به تفصیل شرح داده شده است:
-
تسهیل سنتز رشته لگینگ:
- تکثیر DNA یک فرآیند دوطرفه است، به طوری که یک رشته (رشته پیشرو) به صورت پیوسته سنتز میشود و رشته دیگر (رشته لگینگ) به صورت غیرپیوسته سنتز میشود. قطعات اوکازاکی بخش جداییناپذیر از سنتز رشته لگینگ هستند. آنها به DNA پلیمراز این امکان را میدهند که رشته لگینگ را در جهت 5′ به 3′ سنتز کند، حتی اگر جهتگیری طبیعی آن مخالف جهت تکثیر باشد.
-
جایگزینی RNA پرایمرها با DNA:
- آغاز هر قطعه اوکازاکی با یک RNA پرایمر آغاز میشود. DNA پلیمراز I، نوع خاصی از DNA پلیمراز، مسئول حذف این RNA پرایمرها و جایگزینی آنها با DNA است. این عمل اطمینان میدهد که رشته نهایی تکثیر شده از نوکلئوتیدهای DNA تشکیل شده است.
-
تشکیل رشتههای DNA پیوسته:
- زمانی که قطعات اوکازاکی سنتز شده و RNA پرایمرها با DNA جایگزین شدند، ضروری است که این قطعات به هم متصل شوند تا یک رشته DNA پیوسته شکل بگیرد. DNA لیگاز این عمل را با بستن شکافها در پشتوانه قند-فسفات قطعات انجام میدهد و در نتیجه دو رشته DNA دختر پیوسته و مشابه به هم تشکیل میشود.
-
حفظ یکپارچگی ژنتیکی:
- پردازش صحیح قطعات اوکازاکی برای حفظ یکپارچگی ژنوم ضروری است. نقص در تشکیل یا پردازش این قطعات میتواند منجر به شکستهای رشته DNA و ناهنجاریهای کروموزومی شود. چنین ناهنجاریهایی میتوانند به صورت تغییرات در ظاهر کروموزومها، تغییرات در تعداد کروموزومها یا حتی تغییرات در تعداد مجموعههای کروموزومی ظاهر شوند.
-
حمایت از تقسیم و رشد سلولی:
- تکثیر DNA که با تشکیل و پردازش صحیح قطعات اوکازاکی تسهیل میشود، پیشنیاز ضروری برای تقسیم سلولی است. در موجودات تکسلولی، این تقسیم سلولی به عنوان روشی برای تولید مثل غیرجنسی عمل میکند. در موجودات چندسلولی، تقسیم سلولی برای رشد، ترمیم و تولید سلولهای جدید ضروری است و برای تولید مثل جنسی اهمیت دارد. اطمینان از اینکه هر دو سلول دختر ماده ژنتیکی یکسانی دریافت کنند، برای ادامه حیات و حفظ اطلاعات ژنتیکی ضروری است.
آنزیمهای دخیل در تشکیل قطعات اوکازاکی
-
پریماز (Primase):
- پریماز مسئول اضافه کردن RNA پرایمرها به رشته لگینگ است تا سنتز قطعات اوکازاکی از 5′ به 3′ تسهیل شود. به دلیل سرعت پایین ساخت RNA پرایمر توسط پریماز نسبت به سرعت سنتز DNA پلیمراز، پریماز به عنوان یک سیگنال توقف موقت عمل میکند. این باعث میشود که پیشرفت فورک تکثیر به طور موقت متوقف شود و از پیشی گرفتن رشته لیدینگ نسبت به رشته لگینگ جلوگیری شود
-
DNA پلیمراز (DNA polymerase δ) δ:
- DNA پلیمراز نقش مهمی در سنتز هر دو رشته لیدینگ (پیشرو) و لگینگ دارد. این سنتز شامل سه فاز است که با دو پلیمراز مختلف انجام میشود: DNA پلیمراز α-پریماز و DNA پلیمراز δ. فرآیند با جابجایی RNA و DNA پرایمر توسط بارگذاری کمپلکس کلمپ آغاز میشود که منجر به اتصال کلمپ لغزنده به DNA میشود. سپس، DNA پلیمراز δ سنتز را ادامه میدهد تا به انتهای 5′ قطعه اوکازاکی قبلی برسد. این آنزیم همچنین فعالیت 5′ فلاپ اندونوکلیاز FEN1/RAD27 را تکمیل کرده و از پردازش صحیح قطعات اوکازاکی اطمینان مییابد.
-
DNA لیگاز I (DNA Ligase I) :
- DNA لیگاز I در اتصال قطعات اوکازاکی در حین سنتز رشته لگینگ، به ویژه پس از جایگزینی RNA پرایمرها با DNA توسط DNA پلیمراز δ، اهمیت زیادی دارد. لایگیشن صحیح برای جلوگیری از شکستهای بالقوه در DNA ضروری است. علاوه بر این، پروتئین PCNA (proliferating cell nuclear antigen) نقش کمکی دارد و به DNA لیگاز I کمک میکند تا قطعات را به هم متصل کند.
-
فلاپ اندونوکلیاز 1 (FEN1):
- FEN1 مسئول پردازش قطعات اوکازاکی است. این آنزیم با DNA پلیمراز همکاری میکند تا RNA پرایمر یک قطعه اوکازاکی را حذف کند و همچنین میتواند 5′ ریبونوکلئوتیدها و فلاپهای 5′ را در حین سنتز رشته لگینگ حذف کند. این فرآیند که به آن ترجمه شکاف (nick translation) گفته میشود، قطعات را برای لایگیشن آماده میکند.
-
Dna اندونوکلئاز 2:
- Dna اندونوکلئاز2، اگرچه از نظر ساختاری خاص نیست، برای برش فلاپهای بلند DNA که توسط FEN1 در طول فرآیند بلوغ قطعات اوکازاکی باقی میمانند، ضروری است. این آنزیم برای حذف بخش RNA آغازگر روی قطعات ضروری است. علاوه بر این، Dna2 در متابولیسمهای مختلف DNA و نگهداری تلومرها نقش دارد. فعالیت آن به ویژه زمانی مهم است که بخشهای RNA در انتهای 5′ اتصال یابند، زیرا این آنزیم در جهت 5′ به 3′ عمل میکند. در حضور پروتئین RPA (Single-Stranded DNA-Binding Protein)، زمانی که فلاپهای 5′ DNA کشیده میشوند، Dna2 انتهای 3′ این قطعات را کاهش میدهد و این امکان را فراهم میآورد که FEN1 به طور مؤثر فلاپها را پردازش کند.
در نهایت، تشکیل قطعات اوکازاکی در طول تکثیر DNA یک فرآیند پیچیده است که شامل چندین آنزیم است. هر آنزیم نقش خاص و حیاتی در اطمینان از سنتز دقیق و کارآمد رشته لگینگ ایفا میکند و از یکپارچگی ژنوم محافظت میکند.
آنزیمی که قطعات اوکازاکی را به هم متصل میکند
آنزیمی که قطعات اوکازاکی را به هم متصل میکند، DNA لیگاز است. این آنزیم شکافها بین قطعات اوکازاکی را میبندد و یک رشته DNA پیوسته بر روی رشته لگینگ در حین تکثیر DNA ایجاد میکند.
مکانیسم پیوستگی قطعات اوکازاکی توسط DNA لیگاز:
-
شناسایی شکاف (Nick):
- در حین تکثیر DNA، رشته لگینگ به صورت غیرپیوسته به عنوان قطعات اوکازاکی سنتز میشود. هر قطعه با یک RNA پرایمر آغاز میشود که بعداً با DNA جایگزین میشود. این فرآیند یک شکاف به وجود میآورد که در آن یک شکست در پشتوانه قند-فسفات DNA وجود دارد، به ویژه بین گروه 3′-OH یک نوکلئوتید و گروه 5′-فسفات نوکلئوتید بعدی.
-
فعالسازی DNA لیگاز:
- DNA لیگاز ابتدا با ATP (در یوکاریوتها) یا NAD+ (در برخی پروکاریوتها) فعال میشود. این باعث تشکیل یک پیوند کووالانسی بین یک رزیدوی لیزین در لیگاز و قسمت AMP از ATP میشود که پیروفوسفات آزاد میکند. اکنون این آنزیم “شارژ” شده و آماده عمل است.
-
تشکیل پیوند فسفودی استر:
- آنزیم فعال شده لیگاز سپس AMP را به گروه 5′-فسفات در محل شکاف منتقل میکند. این فرآیند یک واسطه واکنشی ایجاد میکند که در آن DNA آدنیتات میشود.
- گروه 3′-OH نوکلئوتید مجاور شکاف سپس به گروه فسفات آدنیتات حمله میکند و منجر به تشکیل یک پیوند فسفودی استر میشود که شکاف را میبندد.
- در این فرآیند، AMP آزاد میشود.
-
کامل شدن:
- اکنون شکاف در پشتوانه DNA بسته شده و DNA لیگاز آماده است تا بر روی شکاف بعدی عمل کند.
عمل DNA لیگاز تضمین میکند که رشته لگینگ، پس از آن که به صورت قطعات اوکازاکی سنتز شد، به یک رشته DNA پیوسته تبدیل شود که با سنتز پیوستهای که در رشته پیشرو مشاهده میشود، همراستا باشد.
تفاوتهای قطعات اوکازاکی در پروکاریوتها و یوکاریوتها
-
مبداهای تکثیر:
- DNA پروکاریوتها معمولاً یک مبدا تکثیر دارند، در حالی که مولکولهای DNA یوکاریوتها بزرگتر هستند و معمولاً دارای چندین مبدا تکثیر میباشند. این امر منجر به آن میشود که هر کروموزوم یوکاریوتی از واحدهای تکثیر متعدد DNA تشکیل شود، در حالی که در پروکاریوتها فقط یک واحد تکثیر وجود دارد.
-
محیط تکثیر:
- در پروکاریوتها، تکثیر در سیتوپلاسم رخ میدهد. در مقابل، تکثیر DNA در یوکاریوتها در هسته سلول اتفاق میافتد.
-
کمپلکس بارگذاری کلمپ و کلمپها:
- یوکاریوتها یک کمپلکس بارگذاری کلمپ و یک کلمپ شش واحدی به نام پروتئین آنتیژن هستهای سلول تکثیر شونده (PCNA) دارند. این حرکت کارآمد فورک تکثیر اطمینان میدهد که تولید قطعات اوکازاکی با سنتز پیوسته DNA در رشته پیشرو همزمان باشد. کمپلکسهای بارگذاری کلمپ ویژگی خاص یوکاریوتها هستند و تفاوتهای دقیقی را در مقایسه با تکثیر قطعات اوکازاکی در پروکاریوتها نشان میدهند.
-
طول قطعات اوکازاکی:
- قطعات اوکازاکی پروکاریوتی، بهویژه در E. coli، میتوانند تا 2,000 نوکلئوتید طول داشته باشند. در حالی که قطعات اوکازاکی در یوکاریوتها معمولاً کوتاهتر هستند و بین 100 تا 200 نوکلئوتید طول دارند.
-
سرعت تکثیر:
- تکثیر در پروکاریوتها معمولاً سریعتر از یوکاریوتها است. برای مثال، تکثیر باکتریایی میتواند در کمتر از 40 دقیقه تکمیل شود، در حالی که تکثیر سلولهای یوکاریوتی ممکن است تا 400 ساعت طول بکشد.
-
ساختار کروموزومی:
- پروکاریوتها کروموزومهای دایرهای شکل دارند، به همین دلیل فاقد انتهای کروموزومی برای تکثیر هستند. از طرفی، یوکاریوتها کروموزومهای خطی دارند و مکانیسم منحصر به فردی برای تکثیر تلومرهای در انتهای آنها توسعه دادهاند.
-
زمانبندی تکثیر:
- تکثیر در پروکاریوتها فرآیندی مداوم است، در حالی که سلولهای یوکاریوتی تنها در فاز S از چرخه سلولی به تکثیر DNA میپردازند.
-
مقدار DNA:
- سلولهای یوکاریوتی معمولاً حدود 25 برابر DNA بیشتری نسبت به سلولهای پروکاریوتی دارند.
در نهایت، در حالی که هم پروکاریوتها و هم یوکاریوتها از قطعات اوکازاکی در فرآیند تکثیر DNA استفاده میکنند، مکانیسمها، ساختارها و پیچیدگیهای تشکیل و پردازش آنها در دو حوزه مختلف حیات به طور قابل توجهی متفاوت است. این تفاوتها پیچیدگیها و سازگاریهای تکاملی را که در پاسخ به ساختارهای سلولی و چرخههای حیات متفاوت پروکاریوتها و یوکاریوتها به وجود آمدهاند، نشان میدهند.
مقایسه ویژگیها: پروکاریوتها vs. یوکاریوتها (قطعات اوکازاکی)
| ویژگی | پروکاریوتها | یوکاریوتها |
| مبداهای تکثیر | یک مبدا تکثیر واحد. | چندین مبدا تکثیر که منجر به واحدهای متعدد تکثیر دیانای در هر کروموزوم میشود. |
| محیط تکثیر | در سیتوپلاسم رخ میدهد. | در هسته سلول انجام میشود. |
| کلاמפ لودر کمپلکس | غایب. | موجود است و شامل یک کلاپ شش واحدی به نام آنتیژن هستهای در حال تکثیر (PCNA). |
| طول قطعات اوکازاکی | قطعات بلندتر، تا ۲,۰۰۰ نوکلئوتید در E. coli. | قطعات کوتاهتر، معمولاً بین ۱۰۰ تا ۲۰۰ نوکلئوتید. |
| سرعت تکثیر | سریعتر، میتواند در حدود ۴۰ دقیقه تکمیل شود. | کندتر، ممکن است تا ۴۰۰ ساعت در سلولهای حیوانی طول بکشد. |
| ساختار کروموزومی | کروموزومهای دایرهای، فاقد انتهای کروموزومی برای تکثیر. | کروموزومهای خطی با مکانیزمهای خاص برای تکثیر تلومرها در انتهای آنها. |
| زمانبندی تکثیر | تکثیر پیوسته. | تکثیر دیانای فقط در فاز S چرخه سلولی اتفاق میافتد. |
| مقدار دیانای | DNA بسیار کمتری دارد، معمولاً ۲۵ برابر کمتر از سلولهای یوکاریوتی. | دیانای حدوداً ۲۵ برابر بیشتر از سلولهای پروکاریوتی دارد. |
این جدول تفاوتهای اصلی بین پروکاریوتها و یوکاریوتها را در زمینه تکثیر دیانای و فرآیندهای مربوط به قطعات اوکازاکی نشان میدهد. این تفاوتها پیچیدگیهای تکاملی و سازگاریهای منحصر به فرد هر حوزه حیات را بازتاب میدهد.
سوالات متداول
قطعات اوکازاکی چیستند؟
قطعات اوکازاکی توالیهای کوتاه نوکلئوتیدی دیانای هستند که به صورت غیر پیوسته در رشته لگینگ در حین تکثیر دیانای ساخته میشوند.
عملکرد قطعات اوکازاکی چیست؟
عملکرد قطعات اوکازاکی این است که امکان سنتز رشته لگینگ دیانای را در جهت ۵′ به ۳′ فراهم میکنند، که با جهتدار بودن آنزیم دیانای پلیمراز همخوانی دارد.
قطعات اوکازاکی چیستند و چرا تشکیل میشوند؟
قطعات اوکازاکی توالیهای کوتاه دیانای هستند که در رشته لگینگ در حین تکثیر دیانای تشکیل میشوند. این قطعات به دلیل ویژگی ضدپارالل بودن دیانای و جهتدار بودن آنزیم دیانای پلیمراز که تنها قادر به سنتز دیانای در جهت ۵′ به ۳′ است، تشکیل میشوند.
چرا قطعات اوکازاکی تشکیل میشوند؟
آنها برای تسهیل سنتز رشته لگینگ دیانای در جهت ۵′ به ۳′ در حین تکثیر دیانای تشکیل میشوند.
قطعات اوکازاکی در کجا یافت میشوند؟
قطعات اوکازاکی در رشته لگینگ دیانای در حین تکثیر یافت میشوند.
چرا قطعات اوکازاکی غیر پیوسته هستند؟
قطعات اوکازاکی غیر پیوسته هستند زیرا رشته لگینگ به صورت بخشهای کوتاه در جهت مخالف پیشروی شاخه تکثیر سنتز میشود.
کدام قطعه اوکازاکی اول ساخته میشود؟
قطعه اوکازاکی که به مبدا تکثیر نزدیکتر است، اول ساخته میشود.
قطعات اوکازاکی در رشد زنجیره دیانای چیستند؟
در رشد زنجیره دیانای، قطعات اوکازاکی نمایانگر بخشهای کوتاه و غیر پیوسته دیانای هستند که در رشته لگینگ سنتز میشوند.
آیا قطعه اوکازاکی دیانای است یا آرانای؟
قطعه اوکازاکی دیانای است، اما ابتدا با یک پرایمر آرانای آغاز میشود که بعداً با دیانای جایگزین میشود.
همچنین بخوانید:
- همانندسازی نیمه حفاظتی DNA در پروکاریوتها و یوکاریوتها
- همانندسازی DNA-دوبله ژنیران
- همانندسازی ژن چیست؟
- DNA پلیمراز چیست؟
- آسیب DNA و ترمیم DNA: انواع و مکانیسم
مترجم: محمد صادق محمودی لرد