


اطلاعات عمومی
تبادل مواد ژنتیکی بین کروماتیدهای غیر خواهر
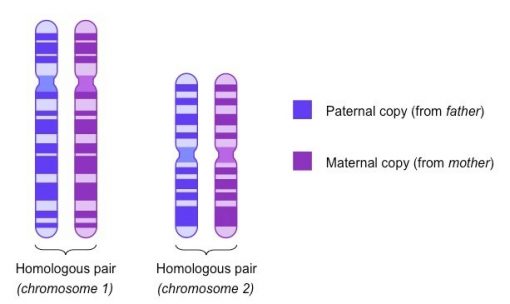
عبور از تبادل مواد ژنتیکی بین کروماتیدهای غیر خواهر کروموزومهای همولوگ در طی میوز انجام میشود که منجر به ترکیب آللی جدید در سلولهای دختر میشود.
هر سلول دیپلوئید شامل دو نسخه از هر کروموزوم است، یکی از گامت مادری و دیگری از گامت پدری. به این جفت کروموزوم که هریک از یک والد مشتق شدهاند، کروموزوم همولوگ میگویند. هنگامی که موجودات دیپلوئید تحت باروری جنسی قرار میگیرند، ابتدا گامت هاپلوئید را از طریق میوز تولید میکنند. در طول پروفاز 1 میوز، کروموزومهای همولوگ با یکدیگر هم راستا شده و مواد ژنتیکی را مبادله میکنند، به طوری که برخی از کروموزومهای حاصل نوترکیب هستند یعنی حاوی ترکیبی از ژنهای کروموزومهای مادری و پدری.
اکثر موجوداتی که تحت باروری جنسی قرار میگیرند دارای دو نوع سلول در بدن خود هستند، سلولهای هاپلوئید و دیپلوئید. سلولهای دیپلوئید در اکثر نقاط بدن دیده میشوند و حاوی دو نسخه از هر کروموزوم هستند. بنابراین، آنها حاوی دو ژن برای تعیین توالی تقریباً هر RNA یا پروتئین هستند. سلولهای هاپلوئید معمولاً گامتهای نر یا ماده هستند که در نتیجه میوز ایجاد شده و در اندامهای جنسی دیده میشوند. آنها فقط یک مجموعه کامل کروموزوم دارند و طوری طراحی شدهاند که با گامت هاپلوئید دیگر ترکیب شوند و یک زیگوت دیپلوئید با یک کروموزوم پدری و یک مجموعه کروموزومی تولید کنند. اگرچه آنها ژنهای یکسانی را کد میکنند، اما توالی DNA آنها میتواند کمی متفاوت از یکدیگر باشد. به عنوان مثال، کروموزوم پدری ممکن است حاوی اطلاعات ژنتیکی باشد که منجر به وجود آنتی ژن A در گلبول های قرمز خون میشود، در حالی که کروموزوم مادر ممکن است آنتی ژن B. را کد کند. این دو آنتی ژن از نظر عملکرد مشابه هستند، اما از نظر دنباله آمینو اسید تا حدودی با یکدیگر تفاوت دارند.
همه سلولهای یک موجود زنده بالغ از زیگوت اولیه مشتق شدهاند و بنابراین از نظر ژنتیکی با یکدیگر یکسان هستند. وقتی این فرد بالغ برای تولید مثل آماده میشود، برخی از سلولهای تخصصی تحت میوز قرار میگیرند و چهار سلول دختر هاپلوئید تولید میکنند. این سلولهای دختر میتوانند پس از بلوغ، گامتهای نر یا ماده را تشکیل دهند.
در طول میوز، کروموزومهای مشتق شده از گامت مادر و پدر میتوانند مستقل از یکدیگر ترکیب شوند. به عبارت دیگر، کروموزومهای مادر یا پدر را میتوان به طور تصادفی برای هر گامت انتخاب کرد. اسپرم انسان میتواند حاوی کروموزومهای 1 ، 3 ، 5 ، 7 ، 9 ، 21 و 23 از والدین زن خود باشد و مابقی میتواند از والد مرد ایجاد شود. به طور مشابه، یک تخمک میتواند فقط دو کروموزوم از والدین زن و مابقی از والدین مرد داشته باشد.
علاوه بر این، هر یک از این کروموزومها میتوانند مخلوطی نوترکیب از ژنهای ناشی از دو والد باشند. یکی از چهار سلول دختر هاپلوئید ناشی از میوز میتواند یک کروموزوم داشته باشد که 80 درصد دنباله آن از والد مرد و 20 درصد باقی مانده از والد زن است. به همین ترتیب، یک سلول دختر دیگر با نسبت معکوس وجود خواهد داشت. این امر از طریق عبور از یکدیگر رخ میدهد، هنگامی که کروموزومهای همولوگ تکراری در نزدیک یکدیگر قرار میگیرند و بخشهایی را با یکدیگر تبادل میکنند.
این تصویر نمایانگر مجموعهای از کروموزومهای همولوگ است. دو مورد از چهار سلول دختر ایجاد شده پس از این رویداد دارای کروموزوم نوترکیبی هستند که نه به طور کامل از مادر گرفته شده است و نه از پدر. این تصویر همچنین نشان میدهد که ژنهایی که در کروموزوم از نظر فیزیکی بسیار نزدیک به یکدیگر هستند به احتمال زیاد با هم به ارث میرسند، در حالی که ژنهایی که در دورتر قرار دارند ممکن است در طول میوز به طور مستقل مرتب شوند.
نقشههای پیوند برآورد فاصله بین دو مکان ژنتیکی، بر اساس فراوانی ترکیب مجدد است. مکانهای ژنی که در یک کروموزوم یکسان از یکدیگر فاصله دارند یا روی کروموزومهای مختلف قرار گرفتهاند، 50 درصد احتمال دارد که با هم به ارث برده شوند و 50 درصد احتمال دارد که به طور مستقل از هم جدا شوند. بر اساس تعداد دفعاتی که آنها با هم به ارث میرسند، می توان نقشههای پیوندی را ایجاد کرد، جایی که موقعیت این ژنها در رابطه با نشانگرهای ژنتیکی شناخته شده نشان داده شده است. به عنوان مثال، اگر یک نشانگر ژنتیکی روی کروموزوم X اغلب همراه با یک آلل خاص برای طاسی الگوی مردانه به ارث برده شود، گفته میشود که آنها بخشی از همان گروه پیوندی هستند. قبل از تعیین توالی DNA با توان بالا، نقشههای پیوند ابزار اصلی برای درک اینکه کدام ژنها در یک کروموزوم خاص وجود دارند، بودند. بعید است که دو نشانگر ژنتیکی که از نظر فیزیکی به یکدیگر نزدیک هستند در طول کراس اوور کروموزومی روی کروماتیدهای مختلف جدا شوند و بنابراین گفته میشود که بیشتر از نشانگرهایی که فاصله زیادی با هم دارند، به هم متصل هستند. به عبارت دیگر، هر دو ژن نزدیکتر بر روی کروموزوم قرار دارند، احتمال ترکیب مجدد بین آنها کمتر است و احتمال به ارث بردن آنها در کنار هم بیشتر است. نشانگرهای کروموزومهای مختلف کاملاً از هم جدا شده اند.
پروفاز I
میوز در دو مرحله رخ میدهد – میوز I و II. میوز I ، مجموعهای از رویدادها است که منجر به تشکیل دو سلول دختر هاپلوئید میشود. در پایان تقسیم کاهشی، تعداد کروموزومها نصف میشود و هر یک از سلولهای دختر تنها یک مجموعه کامل از کروموزومهای کپی شده دارند.
در طول میوز I ، به ویژه پروفاز I ، تعدادی رویداد رخ میدهد، که آن را به یکی از طولانی ترین مراحل میوز تبدیل میکند. این شروع تراکم کروموزوم را نشان میدهد که در آن کروموزومهای کپی شده با کروماتیدهای خواهر متصل، در ابتدا به صورت نخهای نازک بلند دیده میشوند. با تراکم، کروموزومهای همولوگ به دلیل شباهت در ساختار و موقعیت سانترومر کنار هم قرار میگیرند. ساختار پروتئینی به نام کمپلکس سیناپتونمال نیز نقش مهمی ایفا میکند. در این مرحله، کروموزومها به پوشش هستهای متصل میشوند. در حال حاضر، ترکیب مجدد بین کروماتیدهای غیر خواهر کروموزومهای همولوگ اتفاق میافتد. این امر به صورت میکروسکوپی به عنوان عبور از رویداد بین کروموزومهای دو ظرفیتی (یک جفت دو کروموزوم) با ساختار تتراد (کروماتیدهای خواهر تکراری آنها نیز قابل مشاهده است) مشاهده میشود. در اواخر پروفاز I ، به نظر میرسد کروموزومهای همولوگ یکدیگر را “دفع” میکنند. پاکت هستهای دیگر به وضوح قابل مشاهده نیست و سپس سلول به سمت متافاز و آنافاز حرکت میکند تا اولین مرحله میوز را تکمیل کند.
مکانیسم کراسینگ اور
در سطح مولکولی، عبور از یکدیگر با شکستن دو رشته در یکی از مولکولهای DNA آغاز میشود. این شکست دو رشتهای میتواند به طور طبیعی از طریق عواملی مانند اشعه سرطان زا یا از طریق پروتئینهای خاص رخ دهد. متعاقباً، اگزونوکلئازها، آنزیم هایی که نوکلئوتیدها را از انتهای 5 ‘DNA حذف میکنند، در این شکاف عمل کرده و امتداد کوتاه نوکلئوتیدها را در جهت 5’ به 3 ‘از هر دو رشته حذف میکنند. این منجر به آویزان شدن دو ناحیه تک رشتهای میشود که با پروتئینهایی که نوترکیب را کاتالیز میکنند، پوشانده میشوند، همچنین به عنوان نوترکیب شناخته میشوند.این آنزیمها حمله مناطق تک رشتهای به دنبالههایی که برای جفت شدن پایه مناسب هستند، کاتالیز میکنند. نزدیکی کروماتیدهای غیر خواهری در طول پروفاز I ، به این ناحیه تک رشته اجازه میدهد تا از دنباله روی کروموزوم همولوگ استفاده کند. اولین رشته مهاجم مانند یک آغازگر رفتار میکند و یک منطقه دو رشتهای را برای خود با استفاده از یک رشته کروماتید غیر خواهری خود به عنوان الگو ایجاد میکند. این منجر به جابجایی رشته مکمل آن و جفت شدن پایه با دومین ناحیه تک رشتهای میشود که در ابتدا توسط اگزونوکلئاز ایجاد شد. در نهایت، این منجر به تبادل دو رشته با تشکیل یک ساختار متقاطع به نام اتصال هالیدی میشود. این نام از دانشمندی گرفته شده است که برای اولین بار پیشنهاد کرد که چنین پیوندی میتواند هم عبور و هم پدیده دیگری به نام تبدیل ژن را توضیح دهد که در آن محل ژن هتروزیگوت در حین تقسیم سلولی هموزیگوت میشود. اتصالات هالیدی را میتوان از نظر میکروسکوپی به عنوان “کیاسما” در انتهای پروفاز I مشاهده کرد، که تا پایان آنافاز I قابل مشاهده است. اتصالات هالیدی از طریق پروتئینهایی که دستکاری ژنومی را تعدیل میکنند تثبیت شده و به عنوان MSH4 و MSH5 شناخته میشوند.
فواید کراسینگ اور
ارگانیسمهایی که بدون احتمال تجزیه مجدد به صورت غیرجنسی تقسیم میشوند از وضعیتی به نام Ratchet مولر رنج میبرند. یعنی هر نسل از آن گونهها حداقل به اندازه نسل قبلی، اگر نه بیشتر، دارای جهشهای ژنتیکی هستند. به عبارت دیگر، وقتی همه فرزندان از نظر ژنتیکی با یکدیگر یکسان هستند، هیچ راهی برای اصلاح خطاهای ژنتیکی یا ایجاد ترکیبات جدید و مفید وجود ندارد.
کراسینگ اور، تنوع جمعیت را افزایش میدهد و از تجمع ترکیبات مضر آلل جلوگیری میکند، در حالی که اجازه میدهد برخی از ترکیبات والدین به فرزندان منتقل شود. به این ترتیب، بین حفظ ترکیبات آللی مفید و همچنین ایجاد فرصت برای تنوع و تغییر، تعادل وجود دارد.
مترجم: غزل زارعی
مطالعه صدها مطلب علمی در حوزه بیولوژی
آرشیو جدیدترین خبرهای روز دنیای بیولوژی
چرا کراسینک اور نمیتواند همزمان دو تا جایگاه ژنی با هم جابه جا شن
منظورم اینه اگهABCوabcداشته باشیم و کراس هم در bcو BC رخ دهد Abcنداری؟
پاسخش به محدودیت فیزیکی و احتمال وقوع کراسینگاور چندگانه در یک فاصله خاص برمیگردد.
بیایید دقیقتر بررسی کنیم:
فرض:
ژنوتیپ والدها:
کروموزوم اول: A – B – C
کروموزوم دوم (همولوگ): a – b – c
اگر بخوای همزمان بین دو نقطه (یعنی بین B–C و A–B) کراسینگاور بشه، در واقع باید دو کراسینگاور پشتسر هم اتفاق بیفته.
اما چرا همزمان دو کراس اتفاق نمیافته؟
کراسینگاور فرآیندی تصادفی و ناحیهایه:
احتمال اینکه در یک بخش کوچک از کروموزوم، دو کراس همزمان اتفاق بیفته، خیلی کمه. چون بعد از اولین کراس، پروتئینهای دخیل مثل Spo11، Mlh1 و ساختار synaptonemal complex تمایل دارند که اجازه کراس دوم در همون ناحیه رو ندن.
در کراسینگاور معمولاً فقط یک جابهجایی بین دو کروموزوم انجام میشه. اگر بخوای هم A–B و هم B–C کراس کنند، باید دو کراس مستقل، در یک فاصله نزدیک رخ بده. این حالت نادر ولی ممکنه و بهش میگن double crossover.
در double crossover، احتمال تولید Abc کمتر از بقیه گامتهاست چون ترکیب خاصیه که نیاز به دو نقطه شکست داره. بنابراین:
گامت Abc بسیار نادره
بیشتر گامتها یا ABC، abc، یا حاصل از یک کراس منفرد هستن (مثلاً AbC یا aBc)
نتیجه
بهصورت طبیعی، Abc تولید میشه ولی خیلی نادره و به شرطی که دو کراس همزمان بین A–B و B–C رخ بده. به همین دلیل در محاسبات ژنتیکی، همیشه احتمال double crossover رو کمتر در نظر میگیرند و Abc جزء گامتهای غالب نیست.