


اطلاعات عمومی,ویکی ژن
اپران لک چیست؟ تعریف، ساختار، اجزا
فهرست مطالب
نمایش
تعریف اپران لک
- اپران لک یا لاکتوز در اشرشیاکلای، نوعی ژن خوشهای و حاوی سه ژن است که پروتئینهای دخیل در متابولیسم لاکتوز و جایگاههای تنظیمی اپران روی DNA را کد میکنند.
- در باکتریها بسیاری از ژنهای کدکننده پروتئین در اپرانهای واحد رونویسی (که به طور هماهنگ تنظیم میشوند)، کنار هم قرار میگیرند.
- جیکوب و مونود در سال 1961 مدل اپران را برای تنظیم رونویسی پیشنهاد کردند.
اجزای مدل اپران:
- مجموعه ای از ژن های ساختاری (یعنی ژنهای کد کنندهی پروتئینهایی که باید تنظیم شوند).
- یک مکان اوپراتور، که توالیای از DNA است که رونویسی ژنهای ساختاری را تنظیم میکند.
- یک ژن تنظیم کننده که پروتئینی را کد میکند که توالی اوپراتور را تشخیص می دهد.
اپران لک
- یکی از اپرانهای مورد مطالعه، اپران لک در E.coli است.
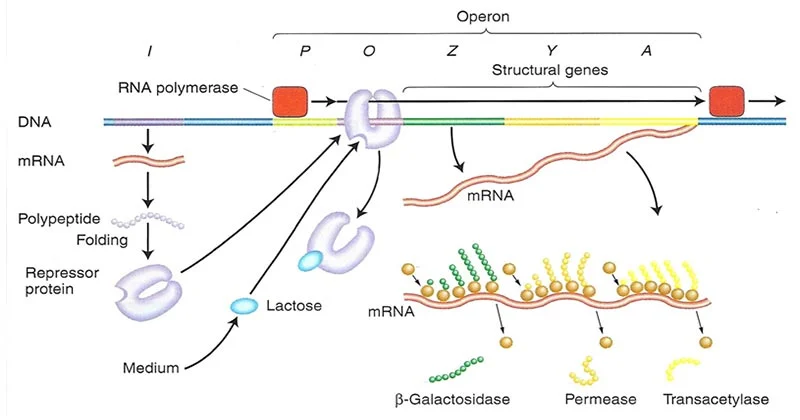
آنزیمهای کلیدی در متابولیسم لاکتوز:
-
گالاکتوزید پرمئاز (همچنین به نام لاکتوز پرمئاز نیز شناخته میشود).
لاکتوز را از طریق غشای سلولی به داخل سلول منتقل میکند.
-
گالاکتوزیداز
لاکتوز را به گلوکز و گالاکتوز هیدرولیز میکند.
-
تیوگالاکتوزید ترانس استیلاز
ساختار اپران لک
- در اپران lac، ژنهای ساختاری ژنهای lacZ، lacY و lacA هستند که به ترتیب گالاکتوزیداز، پرمئاز و ترانس استیلاز را کد میکنند.
- رونویسی از یک پروموتر (Plac) که در بالادست این ژنهای ساختاری قرار دارد و به RNA پلیمراز متصل میشود، رخ میدهد.
با این وجود، یک اوپراتور (Olac) بین پروموتر و ژنهای ساختاری و یک ژن lacI که پروتئین مهارکننده lac را کد میکند، وجود دارد.
- ژن lacI دارای پروموتر خاص خود (PlacI) است که به RNA پلیمراز متصل میشود و منجر به رونویسی mRNA مهارکننده lac و در نتیجه تولید مونومرهای پروتئین مهار کننده lac میشود.
- چهار مونومر مهارکننده یکسان به هم میرسند تا تترامر فعال را تشکیل دهند که میتواند به طور محکم به محل اپراتور lac، یعنی Olac متصل شود.
القا کنندهها و القای اپران لک
- در حالت عادی، سلولهای E. coli به میزان بسیار کمی از هر یک از این سه پروتئین میسازند، اما زمانی که لاکتوز در دسترس باشد، ساخت هر آنزیم بسیار افزایش مییابد.
- بنابراین هر آنزیم یک آنزیم القایی است و این فرآیند را induction (القاء) مینامند.
- مکانیسم به این صورت است که چند مولکول β-گالاکتوزیداز در سلول قبل از القا، لاکتوز را به آلولاکتوز تبدیل میکند که رونویسی این سه ژن را در اپران لک فعال میکند.
- بنابراین آلولاکتوز یک القاء کننده است.
- یکی دیگر از القاء کنندههای اپران لک، ایزوپروپیل تیوگالاکتوزید (IPTG) است. برخلاف آلولاکتوز، این القا کننده توسط E.coli متابولیزه نمیشود بنابراین فقط برای مطالعات تجربی مربوط به القاء مفید است.
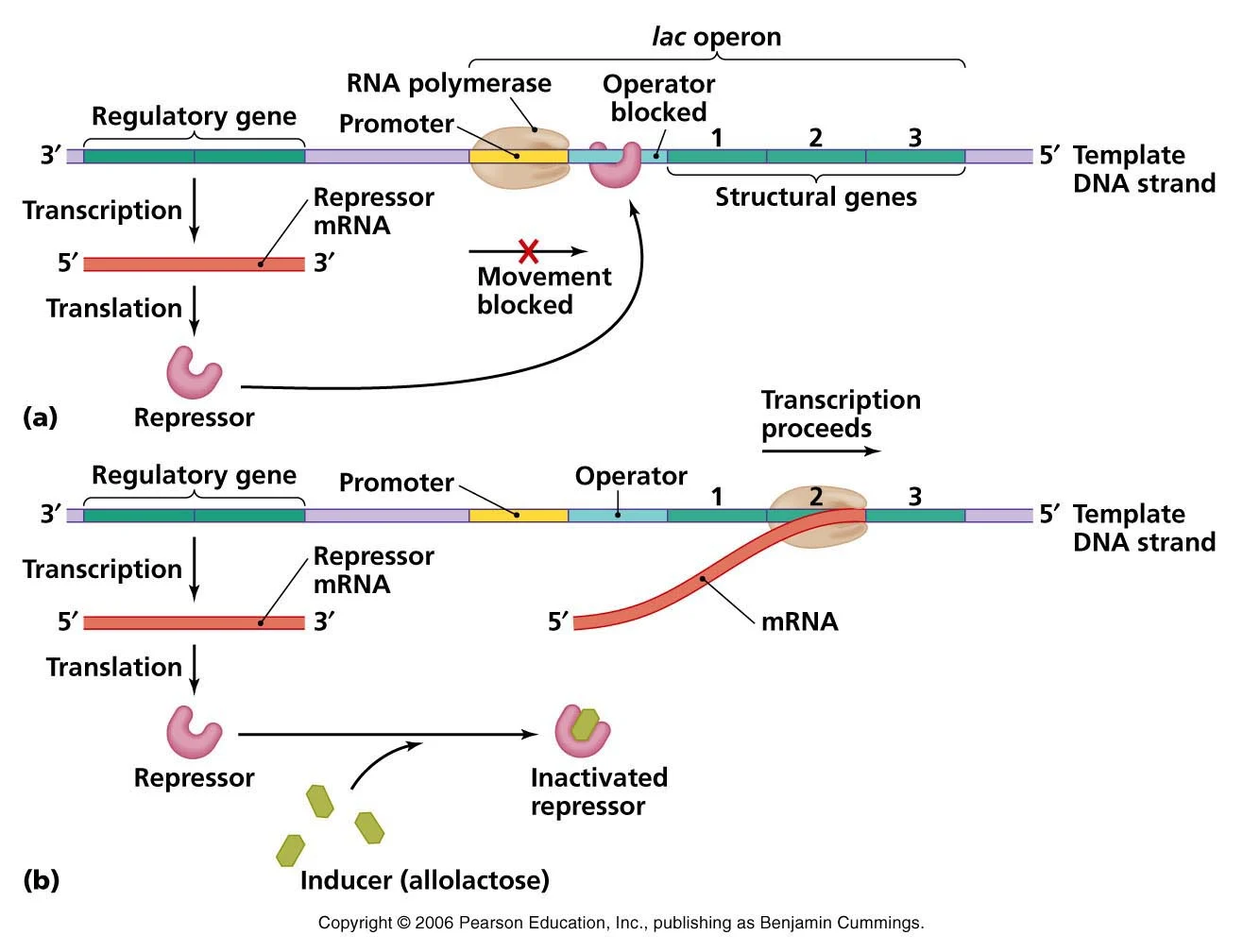
اپران لک در غیاب القاء کننده
- در غیاب القا کنندهای مانند آلولاکتوز یا IPTG، ژن lacI رونویسی میشود و پروتئین مهار کننده حاصل به محل اوپراتور اپران lac، Olac متصل میشود و از رونویسی ژنهای lacZ، lacY و lacA جلوگیری میکند.
اپران لک در حضور القا کنندهها
- در حین القاء، القا کننده به رپرسور یا مهارکننده متصل میشود.
- باعث تغییر در ساختار رپرسور میشود که میل آن را به اتصال به اوپراتور lac بسیار کاهش می دهد.
- رپرسور lac اکنون از محل اوپراتور جدا میشود و به RNA پلیمراز (از قبل در محل پروموتر مجاور وجود دارد) اجازه میدهد تا شروع به رونویسی ژنهای lacZ، lacY و lacA کند.
- ژنها رونویسی میشوند تا یک mRNA پلی سیسترونیک تولید شود که برای تولید هر سه آنزیم در مقادیر زیاد ترجمه میشود.
- وجود mRNA پلی سیسترونیک باعث میشود که مقادیر هر سه محصول ژن، به طور هماهنگ تنظیم شوند.
- اگر القا کننده برداشته شود، مهار کننده لک به سرعت به محل اپراتور لک متصل میشود و رونویسی تقریباً بلافاصله مهار میشود.
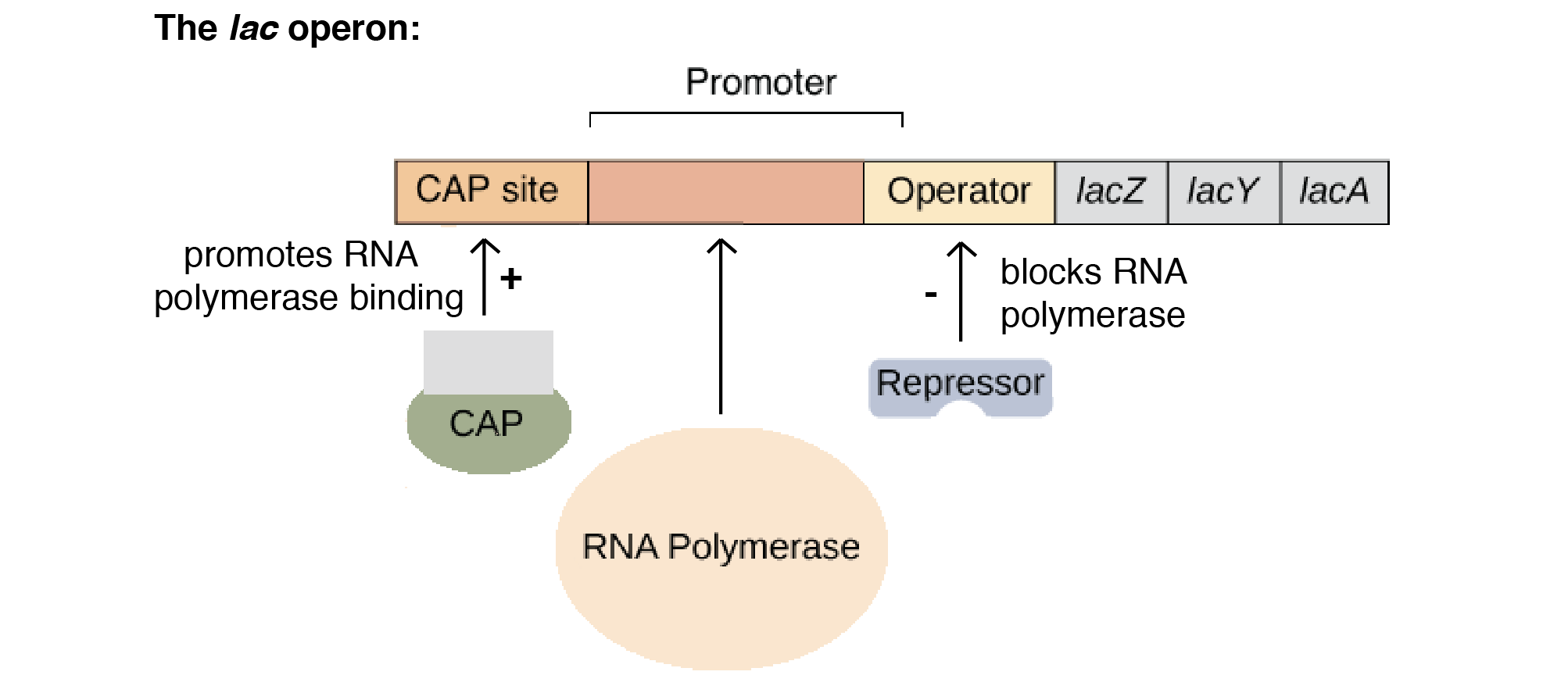
CRP/CAP
- رونویسی در سطح بالای اپران لک، مستلزم وجود یک پروتئین فعال کننده خاص به نام پروتئین فعال کننده کاتابولیت (CAP) است که پروتئین گیرنده cAMP (CRP) نیز نامیده میشود.
- این پروتئین که یک دایمر است نمی تواند به DNA متصل شود مگر اینکه با AMP حلقوی 3ˋ5′ (cAMP) کمپلکس ایجاد کند.
- کمپلکس CRP-cAMP به پروموتر lac درست در بالادست محل اتصال RNA پلیمراز متصل میشود.
- اتصال RNA پلیمراز را افزایش میدهد و بنابراین رونویسی اپران لک را تحریک میکند.
- اینکه آیا پروتئین CRP قادر به اتصال به پروموتر لک است یا نه بستگی به منبع کربن موجود در باکتری دارد.
اپران لک در حضور گلوکز
- هنگامی که گلوکز وجود دارد، E. coli نیازی به استفاده از لاکتوز به عنوان منبع کربن ندارد و بنابراین نیازی نیست که اپران لک فعال باشد.
- بنابراین سیستم تکامل یافته است تا به گلوکز پاسخ دهد.
- گلوکز آدنیلات سیکلاز، آنزیمی که cAMP را از ATP سنتز میکند، مهار میکند.
- بنابراین، در حضور گلوکز، سطح داخل سلولی cAMP کاهش می یابد، بنابراین CRP نمیتواند به پروموتر لک متصل شود و اپران لک در حد ضعیفی فعال است (حتی در حضور لاکتوز).
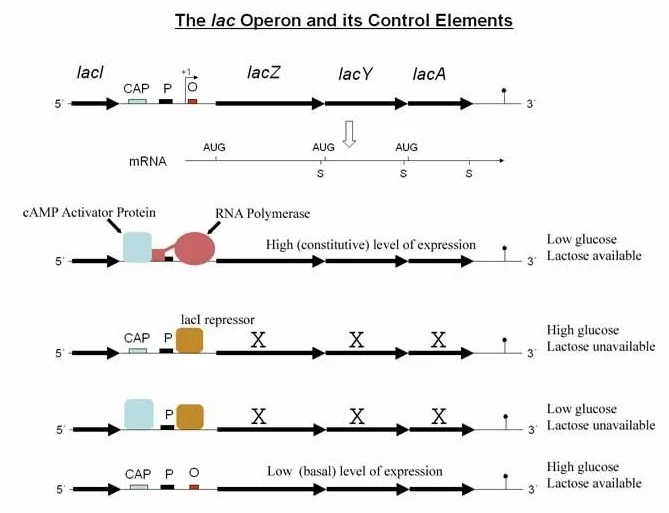
اپران لک در غیاب گلوکز
- هنگامی که گلوکز وجود ندارد، آدنیلات سیکلاز مهار نمیشود، سطح cAMP داخل سلولی افزایش مییابد و به CRP متصل میشود.
- بنابراین، هنگامی که گلوکز وجود ندارد اما لاکتوز وجود دارد، کمپلکس CRP-cAMP رونویسی اپران لک را تحریک میکند و به لاکتوز اجازه میدهد تا به عنوان منبع کربن جایگزین، استفاده شود.
- در غیاب لاکتوز، رپرسور لک، باعث غیر فعال ماندن اپران لک میشود.
- این کنترلهای ترکیبی تضمین میکنند که ژنهای lacZ، lacY و lacA تنها در صورتی که گلوکز وجود نداشته باشد و لاکتوز وجود داشته باشد به میزان زیاد رونویسی میشوند.
تنظیم مثبت و منفی اپران لک
- اپران لک مثال خوبی از کنترل منفی (تنظیم منفی) بیان ژن در آن است که رپرسور متصل از رونویسی ژنهای ساختاری جلوگیری میکند.
- کنترل مثبت یا تنظیم بیان ژن زمانی است که پروتئین تنظیم کننده به DNA متصل میشود و سرعت رونویسی را افزایش میدهد.
- در این حالت پروتئین تنظیم کننده را فعال کننده می نامند. CAP/CRP در تنظیم اپران لک مثال خوبی از یک فعال کننده است.
- بنابراین اپران لک تحت کنترل منفی و مثبت است.
همچنین بخوانید:
- مهندسی ژنتیک چیست؟
- اپرون تریپتوفان
- ترنسداکشن باکتریایی (Bacterial Transduction): تعریف، اصول، مراحل و نمونهها
مترجم: معصومه قریبی ششده
سلام.دراپران لک ،چند کدون آغاز و چند کدون پایان وجود داره؟
در دستگاه ژنتیکی، کدون آغاز برای سنتز پروتئین در تمامی موجودات زنده یکسان است. کدون آغاز معمولاً AUG است که برای آمینواسید متیونین در اوکاریوتها و فرمیل متیونین در پروکاریوتها کد میکند. این کدون نقطه شروع ترجمه را بر روی mRNA مشخص میکند.
در مورد کدونهای پایان، سه کدون وجود دارد که به عنوان سیگنالهای توقف ترجمه عمل میکنند و به پایان ساخت زنجیره پلیپپتیدی منجر میشوند. این سه کدون عبارتند از:
UAA
UAG
UGA
این کدونها در تمامی موجودات از جمله باکتریها، ارکیها، و اوکاریوتها به عنوان کدونهای پایان عمل میکنند و پروتئینسازی را متوقف میکنند. هیچ آمینواسیدی توسط این کدونها کد نمیشود و آنها صرفاً به ریبوزوم دستور میدهند که سنتز پروتئین را متوقف کرده و اجزای ترجمه را آزاد کند.
پس در پاسخ به سوال شما، یک کدون آغاز (AUG) و سه کدون پایان (UAA, UAG, UGA) در دستگاه ژنتیکی وجود دارد.